
Biodiversity of Angola pp 543–549 Cite as

Conclusions: Biodiversity Research and Conservation Opportunities
- Vladimir Russo 5 ,
- Brian J. Huntley 6 ,
- Fernanda Lages 7 &
- Nuno Ferrand 8 , 9 , 10
- Open Access
- First Online: 21 February 2019
19k Accesses
3 Citations
Angola is a country full of opportunities. Few countries offer more exciting prospects for young scientists to discover and document the rich biodiversity, complex ecosystem processes and undescribed species of plants and animals that are to be found in its amazing diversity of landscapes and seascapes. The current expanding support of the Angolan government and of international partners is unprecedented, and the positive response from young students ensures the growth of a new generation of biodiversity researchers and conservation professionals. Based on a synthesis of biodiversity research and conservation activities of the past century, we outline opportunities, approaches and priorities for a strengthened collaborative research and conservation agenda.
You have full access to this open access chapter, Download chapter PDF
Context: Challenges and Opportunities
This book was conceived on the basis of three simplistic assumptions. First, that very little information is available on the biodiversity of Angola. The second assumption was that colonial governments had neglected, and post-independence authorities have been unsupportive of research on the fauna, flora and ecosystems of Angola. Thirdly, we assumed that existing biodiversity knowledge was mostly lost in dusty government archives or inaccessible scientific journals. In truth, the rich assemblage of information gathered together in this synthesis volume demonstrates the error of these assumptions. The perceived challenges of the past have become a mosaic of opportunities for the future.
The energy, knowledge and dedication of this book’s 46 contributing authors has resulted in a comprehensive synopsis of the ‘state of the science’ on the evolution and diversity of Angola’s landscapes, flora, vegetation, all vertebrates, two iconic invertebrate taxa, and key ecosystems in both marine and terrestrial environments. What is immediately obvious from each chapter is the wealth of Angola’s natural heritage, and how fragile it is to anthropogenic impacts and the vicissitudes of climate change. The vulnerability of the remnant forests of Angolan Escarpment and Afromontane ecosystems to these pressures places a tremendous responsibility on Angolans to study and protect these fingerprints of the past. Angola’s universities, scientific organisations and government research institutions can lead the way to strengthening our understanding of the evolution, structure and functioning of these and the many other special habitats that make Angola unique in Africa in terms of the diversity of biomes and ecoregions found within its borders.
Each chapter of this volume reveals research and conservation opportunities relevant to the environment or taxon under discussion and presents compelling arguments for greater levels of investment in both research and conservation. Some needs are very specific, such as the importance of biodiversity surveys, vegetation maps and socio-economic assessments of the country’s many protected areas as a basis to achieving their effective management and the delivery of long-term benefits to society. Other priorities are more broadly based, aimed at developing tools for wide-scale natural resource-use planning, such as a new and detailed vegetation map for the entire country, building on the results of the current forest inventory, and for a national biodiversity data portal. Yet others focus on selected taxa that can help, through modern genomic studies, to explain the processes of speciation that have led to the richness of the country’s flora and fauna. All have a common purpose – to effectively inform decisions that will ensure increasingly sustainable development for all Angolans and for humanity at large.
Towards a Biodiversity Conservation Research Strategy
Over 40 research topics are identified in the chapters of this volume. Such compilations of research opportunities need to be embraced within national strategies, matching needs with resources and priorities within a research agenda, while understanding the country’s challenges. In recent decades, a first point of departure in the design of conservation science programmes has been the frameworks provided by international multilateral environmental conventions – most specifically the Convention on Biological Diversity to which Angola has been a signatory since 1998. The development of Angola’s National Biodiversity Strategy and Action Plan (NBSAP) brought a logical structure to both policy and planning (GoA 2006 , 2018 ). These strategies have been further developed through setting nationally relevant implementation goals such as those detailed in the Aichi Targets (CBD 2010 ). An early lesson learned in the implementation of such strategies was the fundamental importance of cooperative partnerships between multiple institutions. The global scarcity of taxonomists, for example, has meant that no single country has the capacity to study, understand and document all taxa and ecosystems. The strength of organisations such as IUCN, GBIF, IPBES, etc., is based on shared information and skills. The benefits of the recent surge of interest in Angola’s biodiversity demonstrated by foreign universities, museums, non-governmental organisations and intergovernmental agreements have yet to be fully exploited. For this, a strategic approach, implemented opportunistically, is advantageous.
Building a programme of work for conservation science needs to be mobilised at several levels, often simultaneously, rather than sequentially. At a first level, biodiversity assessments are a priority. Biodiversity surveys, which provide both the building blocks of knowledge and unique training and capacity building opportunities, must maintain the positive momentum of the past decade. The preliminary checklists of species presented in this volume should be regularly updated and expanded to integrate these and other taxonomic groups through an electronic biodiversity data portal. Ideally, international institutions, in liaison with national hosts, should assist in the coordination and curation of checklists, atlases and field guidebooks such as those already available for the reptiles and amphibians of Cangandala National Park (Ceríaco et al. 2016 ), the ‘special birds’ of Angola (Mills 2018 ) and the atlas of Angolan reptiles and amphibians (Marques et al. 2018 ). Checklists and the natural history collections on which they are based need integration within international databases such as those of GBIF and the Catalogue of Life. A direct outcome of this component of research is the development of taxonomic skills and of para-taxonomists in Angola.
Second, the socio-ecological systems within which research and conservation play out should not be neglected. Biodiversity does not exist in a vacuum – it has a human face that governs the success or failure of interventions. Biodiversity scientists who ignore the human dimensions of research and conservation do so at their peril. This is especially true within and around formal protected areas, where studies are needed on the socio-economic drivers of change (from a subsistence to a cash-based economy in rural areas), and the impacts of the bushmeat trade, slash-and-burn agricultural practices, charcoal production and nomadic pastoralism, on biodiversity conservation across the country. Such studies should also draw on local indigenous knowledge in developing management plans and research projects. At a regional scale, the assessment of the role of ecosystem services for sustainable living in rural and urban environments provides information essential to long-term development planning, ideally guided by a new and detailed vegetation map of Angola. The interdependence of basic research, biodiversity assessments and applied studies is obvious.
Third, there is no questioning the paramount importance of strengthening the capacity of young Angolan researchers and of research institutions. These actions should be framed within a collaborative and mutually beneficial strategy. International collaboration is already assisting in this, but needs further support. Scholarships, internships and mentoring programmes are fundamental, but field experience is critical for young biodiversity researchers, and the promotion of bush camps for student training in protected areas and biodiversity hotspots is a highly effective mechanism for inspiring the new generation. The establishment of a virtual network of Angolan conservation professionals using social media such as the Angola Ambiente Facebook site is a highly effective initiative. Angolan institutions also need strengthening and closer cooperation across government departments, and the integration of their research results through a formal information portal, is a key opportunity.
Research Opportunities from Genes to Landscapes
While the above three pillars form the foundations of a nascent biodiversity conservation research agenda, with an emphasis on immediate and practical needs, there are many fascinating questions relating to the functioning of Angola’s diverse ecosystems that challenge the minds of biologists as they explore the country. The research strategy should be reinforced by studies on the evolutionary and ecological processes that account for Angola’s biodiversity. An understanding of the evolution of Angola’s biota will be strengthened by the development of modern phylogenies of key taxonomic groups, as proposed in many to the chapters of this volume. An understanding of the evolution of species assemblages and of individual species is of great value to guiding conservation measures, as already demonstrated by the Angolan studies of Vaz da Silva ( 2015 ) and Vaz Pinto ( 2018 ). Advanced molecular technologies allow new insights into many patterns revealed by basic surveys and assessments. The population genetics and hybridisation of Angola’s two elephant species needs urgent study before the last remnants of Forest Elephant are driven to extinction. Similarly, iconic plants such as the baobab Adansonia digitata that in Angola manifests as widely divergent phenotypes, from dwarf baobabs in Namibe to the obese giants of Cunene and the slender columns of Cuanza-Norte, merit studies on their genetic diversity, ecology and traditional uses. Angola’s once vast populations of Welwitschia mirabilis await research on their population dynamics and potential resilience to overgrazing by cattle in Iona National Park. The bizarre patterns of ‘fairy circles’ of the Namib, ‘fairy forests’ of the Lundas and Moxico, of dwarf succulents along the desert margins of Benguela, and the ecological role of fog along the entire coast, are all ecological puzzles that need elucidation.
Many of these topics might at first sight appear of little more than academic interest, but every element of applied science and technology rests on the fundamentals of curiosity-driven enquiry. The baseline compilation of checklists leads to the identification of patterns of endemism and rarity, to be understood through phylogeographic studies across the country. From such studies, increasingly robust scenarios of evolutionary processes may be built. It is these phenomena of the patterns in nature, their ecological functioning and the interactions that drive large-scale environmental dynamics that will ultimately guide sustainable landuse management and inform responses to the impacts of climate change. It is at the level of landscapes and seascapes that the nation’s economy and progress is built, and such wide-horizon visioning, underpinned by fundamental research, is needed for the sustainable development agendas of the twenty-first century.
At a landscape scale, an early priority should be to update the vegetation map of Angola to give a more balanced and objective delineation of the country’s major vegetation units, for landuse planning and conservation purposes. This can best be achieved with the input of a multi-national team of workers, using modern remote sensing technologies for vegetation classification, mapping and monitoring. Vegetation classification and mapping skills take many years, even decades to develop, and the close collaboration, even leadership, of foreign experts would be valuable. An improved vegetation map will help expedite assessments of ecosystem conservation status, landuse potential, research priorities and opportunities, and help develop a predictive understanding of ecosystem structure and function.
At ecosystem scale, the importance of securing the effective management of protected areas, large and small, terrestrial and marine, is a sine qua non to the future of Angola’s biodiversity. The biological and cultural importance of relatively small protected areas, such as Ilhéu dos Pássaros, and future protected areas such as Mount Moco, Namba, Cumbira, Tundavala and the rainforests of Cuanza-Norte, Uíge and Zaire, and of turtle nesting grounds along the coast, should not be overlooked during the pursuit of mega-parks that might excite the public and motivate politicians. Targeted studies of the existing protected areas and of the key biodiversity hotspots identified by the Ministry of Environment in successive strategies (GoA 2006 , 2011 , 2018 ) should be given priority, as these protected areas and hotspots most probably hold more than 80% of Angola’s floral and faunal diversity in less than 15% of the country’s land area. As training grounds for young biologists and conservation scientists, protected areas have no equal. Furthermore, identifying and rigorously protecting near-pristine zones within otherwise threatened protected areas such as Quiçama, Luando, Iona and Mupa, and of the country’s extensive coastline, should be an integral part of any protected areas strategy. The marine environment is especially sensitive to the impacts of human activities and science-based marine spatial planning is essential if long-term conflicts between humans and the marine environment are to be avoided. The importance of a focus on the biodiversity hotspots and on the existing protected areas of Angola – the repositories of the country’s natural wealth – is self-evident.
Conclusions
These outlines are of necessity simplistic and preliminary. Each of the chapters in this book identifies research questions that can be addressed by an emerging generation of Angolan biodiversity scientists and conservation professionals. The challenges are exciting and demanding – offering multiple opportunities for intellectual stimulation, knowledge generation and international collaboration. Angola is truly alive with research and conservation opportunities. The country is still blessed with vast areas of rich wilderness and unique habitats, and has the opportunity to stimulate scientists, conservationists and the general public to participate in programmes of research and effective biodiversity conservation management. As this synthesis volume demonstrates, the limits are boundless. Carpe diem !
CBD (2010) Strategic plan for biodiversity 2011–2020 and the Aichii targets. Secretariat of the Convention on Biological Diversity, Montreal
Google Scholar
Ceríaco LMP, Marques MP, Bandeira SA et al (2016) Anfíbios e Répteis do Parque Nacional da Cangandala. Instituto Nacional da Biodiversidade e Áreas de Conservação/Museu Nacional de História Natural e da Ciência, Luanda/Lisboa, 96 pp
GoA (Government of Angola) (2006) National biodiversity strategy and action plan (2007–2012). Ministry of Urban Affairs and the Environment, Luanda, 55 pp
GoA (Government of Angola) (2011) Plano Estratégico da Rede Nacional de Áreas de Conservação de Angola (PLERNACA). Ministério do Ambiente, Luanda
GoA (Government of Angola) (2018) Plano Estratégico para o Sistema de Áreas de Conservação de Angola (PESAC). Ministério do Ambiente, Luanda
Marques MP, Ceríaco LMP, Blackburn DC, Bauer AM (2018) Diversity and distribution of the amphibians and terrestrial reptiles of Angola. Atlas of historical and bibliographic records (1840–2017). Proceedings of the California Academy of Sciences Series 4, 65:1–501
Mills MSL (2018) The special birds of Angola/As Aves Especiais de Angola. Go-Away-Birding, Cape Town
Vaz da Silva B (2015) Evolutionary history of the birds of the Angolan highlands – the missing piece to understand the biogeography of the Afromontane forests. MSc thesis. University of Porto, Porto
Vaz Pinto P (2018) Evolutionary history of the critically endangered giant sable antelope (Hippotragus niger variani): insights into its phylogeography, population genetics, demography and conservation. PhD thesis. University of Porto, Porto
Download references
Author information
Authors and affiliations.
Fundação Kissama, Luanda, Angola
Vladimir Russo
CIBIO-InBIO, Centro de Investigação em Biodiversidade e Recursos Genéticos, Universidade do Porto, Vairão, Portugal
Brian J. Huntley
ISCED – Instituto Superior de Ciências da Educação da Huíla, Lubango, Angola
Fernanda Lages
CIBIO-InBIO, Centro de Investigação em Biodiversidade e Recursos Genéticos, Laboratório Associado, Campus de Vairão, Universidade do Porto, Vairão, Portugal
Nuno Ferrand
Departamento de Biologia, Faculdade de Ciências, Universidade do Porto, Porto, Portugal
Department of Zoology, Auckland Park, University of Johannesburg, Johannesburg, South Africa
You can also search for this author in PubMed Google Scholar
Corresponding author
Correspondence to Vladimir Russo .
Editor information
Editors and affiliations.
CIBIO-InBIO, Centro de Investigação em Biodiversidade e Recursos Genéticos Laboratório Associado, Campus de Vairão, Universidade do Porto, Vairão, Portugal
Rights and permissions
Open Access This chapter is licensed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license and indicate if changes were made.
The images or other third party material in this chapter are included in the chapter's Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the chapter's Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
Reprints and permissions
Copyright information
© 2019 The Author(s)
About this chapter
Cite this chapter.
Russo, V., Huntley, B.J., Lages, F., Ferrand, N. (2019). Conclusions: Biodiversity Research and Conservation Opportunities. In: Huntley, B., Russo, V., Lages, F., Ferrand, N. (eds) Biodiversity of Angola. Springer, Cham. https://doi.org/10.1007/978-3-030-03083-4_20
Download citation
DOI : https://doi.org/10.1007/978-3-030-03083-4_20
Published : 21 February 2019
Publisher Name : Springer, Cham
Print ISBN : 978-3-030-03082-7
Online ISBN : 978-3-030-03083-4
eBook Packages : Biomedical and Life Sciences Biomedical and Life Sciences (R0)
Share this chapter
Anyone you share the following link with will be able to read this content:
Sorry, a shareable link is not currently available for this article.
Provided by the Springer Nature SharedIt content-sharing initiative
- Publish with us
Policies and ethics
- Find a journal
- Track your research
Thank you for visiting nature.com. You are using a browser version with limited support for CSS. To obtain the best experience, we recommend you use a more up to date browser (or turn off compatibility mode in Internet Explorer). In the meantime, to ensure continued support, we are displaying the site without styles and JavaScript.
- View all journals
- Explore content
- About the journal
- Publish with us
- Sign up for alerts
- Published: 24 August 2020
The results of biodiversity–ecosystem functioning experiments are realistic
- Malte Jochum ORCID: orcid.org/0000-0002-8728-1145 1 , 2 , 3 ,
- Markus Fischer 1 ,
- Forest Isbell ORCID: orcid.org/0000-0001-9689-769X 4 ,
- Christiane Roscher ORCID: orcid.org/0000-0001-9301-7909 2 , 5 ,
- Fons van der Plas ORCID: orcid.org/0000-0003-4680-543X 6 ,
- Steffen Boch ORCID: orcid.org/0000-0003-2814-5343 1 , 7 ,
- Gerhard Boenisch 8 ,
- Nina Buchmann ORCID: orcid.org/0000-0003-0826-2980 9 ,
- Jane A. Catford ORCID: orcid.org/0000-0003-0582-5960 10 ,
- Jeannine Cavender-Bares ORCID: orcid.org/0000-0003-3375-9630 4 ,
- Anne Ebeling 11 ,
- Nico Eisenhauer ORCID: orcid.org/0000-0002-0371-6720 2 , 3 ,
- Gerd Gleixner 8 ,
- Norbert Hölzel 12 ,
- Jens Kattge ORCID: orcid.org/0000-0002-1022-8469 2 , 8 ,
- Valentin H. Klaus ORCID: orcid.org/0000-0002-7469-6800 9 ,
- Till Kleinebecker ORCID: orcid.org/0000-0003-1121-2861 13 ,
- Markus Lange ORCID: orcid.org/0000-0002-2802-9177 8 ,
- Gaëtane Le Provost ORCID: orcid.org/0000-0002-1643-6023 14 ,
- Sebastian T. Meyer ORCID: orcid.org/0000-0003-0833-1472 15 ,
- Rafael Molina-Venegas ORCID: orcid.org/0000-0001-5801-0736 1 , 16 ,
- Liesje Mommer 17 ,
- Yvonne Oelmann ORCID: orcid.org/0000-0003-3513-6568 18 ,
- Caterina Penone ORCID: orcid.org/0000-0002-8170-6659 1 ,
- Daniel Prati 1 ,
- Peter B. Reich ORCID: orcid.org/0000-0003-4424-662X 19 , 20 ,
- Abiel Rindisbacher ORCID: orcid.org/0000-0001-8605-2976 1 ,
- Deborah Schäfer 1 ,
- Stefan Scheu ORCID: orcid.org/0000-0003-4350-9520 21 , 22 ,
- Bernhard Schmid ORCID: orcid.org/0000-0002-8430-3214 23 , 24 ,
- David Tilman ORCID: orcid.org/0000-0001-6395-7676 4 , 25 ,
- Teja Tscharntke 26 ,
- Anja Vogel 2 , 3 , 11 ,
- Cameron Wagg ORCID: orcid.org/0000-0002-9738-6901 27 ,
- Alexandra Weigelt ORCID: orcid.org/0000-0001-6242-603X 2 , 6 ,
- Wolfgang W. Weisser ORCID: orcid.org/0000-0002-2757-8959 15 ,
- Wolfgang Wilcke ORCID: orcid.org/0000-0002-6031-4613 28 &
- Peter Manning ORCID: orcid.org/0000-0002-7940-2023 14
Nature Ecology & Evolution volume 4 , pages 1485–1494 ( 2020 ) Cite this article
8615 Accesses
82 Citations
216 Altmetric
Metrics details
- Biodiversity
- Community ecology
- Grassland ecology
A large body of research shows that biodiversity loss can reduce ecosystem functioning. However, much of the evidence for this relationship is drawn from biodiversity–ecosystem functioning experiments in which biodiversity loss is simulated by randomly assembling communities of varying species diversity, and ecosystem functions are measured. This random assembly has led some ecologists to question the relevance of biodiversity experiments to real-world ecosystems, where community assembly or disassembly may be non-random and influenced by external drivers, such as climate, soil conditions or land use. Here, we compare data from real-world grassland plant communities with data from two of the largest and longest-running grassland biodiversity experiments (the Jena Experiment in Germany and BioDIV in the United States) in terms of their taxonomic, functional and phylogenetic diversity and functional-trait composition. We found that plant communities of biodiversity experiments cover almost all of the multivariate variation of the real-world communities, while also containing community types that are not currently observed in the real world. Moreover, they have greater variance in their compositional features than their real-world counterparts. We then re-analysed a subset of experimental data that included only ecologically realistic communities (that is, those comparable to real-world communities). For 10 out of 12 biodiversity–ecosystem functioning relationships, biodiversity effects did not differ significantly between the full dataset of biodiversity experiments and the ecologically realistic subset of experimental communities. Although we do not provide direct evidence for strong or consistent biodiversity–ecosystem functioning relationships in real-world communities, our results demonstrate that the results of biodiversity experiments are largely insensitive to the exclusion of unrealistic communities and that the conclusions drawn from biodiversity experiments are generally robust.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
24,99 € / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
111,21 € per year
only 9,27 € per issue
Rent or buy this article
Prices vary by article type
Prices may be subject to local taxes which are calculated during checkout
Similar content being viewed by others

Species richness is more important for ecosystem functioning than species turnover along an elevational gradient
Jörg Albrecht, Marcell K. Peters, … Matthias Schleuning

Biodiversity–stability relationships strengthen over time in a long-term grassland experiment
Cameron Wagg, Christiane Roscher, … Bernhard Schmid

Consistent stabilizing effects of plant diversity across spatial scales and climatic gradients
Maowei Liang, Benjamin Baiser, … Shaopeng Wang
Data availability
The data supporting the findings of our study are available at https://doi.org/10.25829/idiv.1869-11-3082 .
Code availability
The R code to reproduce the findings and figures of our study is available at https://doi.org/10.25829/idiv.1869-11-3082 .
Cardinale, B. J. et al. Biodiversity loss and its impact on humanity. Nature 486 , 59–67 (2012).
Article CAS Google Scholar
Tilman, D., Isbell, F. & Cowles, J. M. Biodiversity and ecosystem functioning. Annu. Rev. Ecol. Evol. Syst. 45 , 471–493 (2014).
Article Google Scholar
Isbell, F. et al. Linking the influence and dependence of people on biodiversity across scales. Nature 546 , 65–72 (2017).
van der Plas, F. Biodiversity and ecosystem functioning in naturally assembled communities. Biol. Rev. 94 , 1220–1245 (2019).
Google Scholar
Schulze, E.-D. & Mooney, H. Biodiversity and Ecosystem Functioning (Springer, 1993).
Naeem, S., Thompson, L. J., Lawler, S. P., Lawton, J. H. & Woodfin, R. M. Declining biodiversity can alter the performance of ecosystems. Nature 368 , 734–737 (1994).
Balvanera, P. et al. Quantifying the evidence for biodiversity effects on ecosystem functioning and services. Ecol. Lett. 9 , 1146–1156 (2006).
Hines, J. et al. Mapping change in biodiversity and ecosystem function research: food webs foster integration of experiments and science policy. Adv. Ecol. Res. 61 , 297–322 (2019).
Tilman, D., Wedin, D. & Knops, J. Productivity and sustainability influenced by biodiversity in grassland ecosystems. Nature 379 , 718–720 (1996).
Roscher, C., Schumacher, J. & Baade, J. The role of biodiversity for element cycling and trophic interactions: an experimental approach in a grassland community. Basic Appl. Ecol. 121 , 107–121 (2004).
Tilman, D. et al. Diversity and productivity in a long-term grassland experiment. Science 294 , 843–845 (2001).
Hooper, D. U. et al. Effects of biodiversity on ecosystem functioning: a consensus of current knowledge. Ecol. Monogr. 75 , 3–35 (2005).
Cardinale, B. J. et al. The functional role of producer diversity in ecosystems. Am. J. Bot. 98 , 572–592 (2011).
O’Connor, M. I. et al. A general biodiversity–function relationship is mediated by trophic level. Oikos 126 , 18–31 (2017).
Loreau, M. et al. Biodiversity and ecosystem functioning: current knowledge and future challenges. Science 294 , 804–808 (2001).
Reich, P. B. et al. Impacts of biodiversity loss escalate through time as redundancy fades. Science 336 , 589–592 (2012).
Huston, M. A. Hidden treatments in ecological experiments: re-evaluating the ecosystem function of biodiversity. Oecologia 110 , 449–460 (1997).
Grime, J. P. Benefits of plant diversity to ecosystems: immediate, filter and founder effects. J. Ecol. 86 , 902–910 (1998).
Wardle, D. A. et al. Biodiversity and ecosystem function: an issue in ecology. Bull. Ecol. Soc. Am. 81 , 235–239 (2000).
Leps, J. What do the biodiversity experiments tell us about consequences of plant species loss in the real world? Basic Appl. Ecol. 5 , 529–534 (2004).
Srivastava, D. S. & Vellend, M. Biodiversity–ecosystem function research: is it relevant to conservation? Annu. Rev. Ecol. Evol. Syst. 36 , 267–294 (2005).
Duffy, J. E. Why biodiversity is important to the functioning of real-world ecosystems. Front. Ecol. Environ. 7 , 437–444 (2008).
Duffy, J. E. Biodiversity effects: trends and exceptions—a reply to Wardle and Jonsson. Front. Ecol. Environ. 8 , 11–12 (2010).
Wardle, D. A. & Jonsson, M. Biodiversity effects in real ecosystems—a response to Duffy. Front. Ecol. Environ. 8 , 10–11 (2010).
Wardle, D. A. Do experiments exploring plant diversity–ecosystem functioning relationships inform how biodiversity loss impacts natural ecosystems? J. Veg. Sci. 27 , 646–653 (2016).
Manning, P. et al. Transferring biodiversity-ecosystem function research to the management of ‘real-world’ ecosystems. Adv. Ecol. Res. 61 , 323–356 (2019).
Wilsey, B. J. & Potvin, C. Biodiversity and ecosystem functioning: importance of species evenness in an old field. Ecology 81 , 887–892 (2000).
Wilsey, B. J. & Polley, H. W. Realistically low species evenness does not alter grassland species-richness–productivity relationships. Ecology 85 , 2693–2700 (2004).
Hillebrand, H., Bennett, D. & Cadotte, M. Consequences of dominance: a review of evenness effects on local and regional ecosystem processes. Ecology 89 , 1510–1520 (2008).
Schmitz, M. et al. Consistent effects of biodiversity on ecosystem functioning under varying density and evenness. Folia Geobot. 48 , 335–353 (2013).
Finn, J. A. et al. Ecosystem function enhanced by combining four functional types of plant species in intensively managed grassland mixtures: a 3-year continental-scale field experiment. J. Appl. Ecol. 50 , 365–375 (2013).
Weisser, W. W. et al. Biodiversity effects on ecosystem functioning in a 15-year grassland experiment: patterns, mechanisms, and open questions. Basic Appl. Ecol. 23 , 1–73 (2017).
Schmid, B. & Hector, A. The value of biodiversity experiments. Basic Appl. Ecol. 5 , 535–542 (2004).
Eisenhauer, N. et al. Biodiversity–ecosystem function experiments reveal the mechanisms underlying the consequences of biodiversity change in real world ecosystems. J. Veg. Sci. 27 , 1061–1070 (2016).
Isbell, F. et al. Nutrient enrichment, biodiversity loss, and consequent declines in ecosystem productivity. Proc. Natl Acad. Sci. USA 110 , 11911–11916 (2013).
Duffy, J. E., Godwin, C. M. & Cardinale, B. J. Biodiversity effects in the wild are common and as strong as key drivers of productivity. Nature 549 , 261–264 (2017).
Buchmann, T. et al. Connecting experimental biodiversity research to real-world grasslands. Perspect. Plant Ecol. Evol. Syst. 33 , 78–88 (2018).
Tilman, D. et al. The influence of functional diversity and composition on ecosystem processes. Science 277 , 1300–1302 (1997).
Tilman, D., Reich, P. B. & Isbell, F. Biodiversity impacts ecosystem productivity as much as resources, disturbance, or herbivory. Proc. Natl Acad. Sci. USA 109 , 10394–10397 (2012).
Isbell, F. et al. Biodiversity increases the resistance of ecosystem productivity to climate extremes. Nature 526 , 574–577 (2015).
Fischer, M. et al. Implementing large-scale and long-term functional biodiversity research: the biodiversity exploratories. Basic Appl. Ecol. 11 , 473–485 (2010).
Soliveres, S. et al. Biodiversity at multiple trophic levels is needed for ecosystem multifunctionality. Nature 536 , 456–459 (2016).
Tilman, D. Secondary succession and the pattern of plant dominance along experimental nitrogen gradients. Ecol. Monogr. 57 , 189–214 (1987).
Clark, C. M. & Tilman, D. Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands. Nature 451 , 712–715 (2008).
Inouye, R. et al. Old-field succession on a Minnesota sand plain. Ecology 68 , 12–26 (1987).
Díaz, S. et al. The global spectrum of plant form and function. Nature 529 , 167–171 (2015).
Craven, D. et al. Multiple facets of biodiversity drive the diversity–stability relationship. Nat. Ecol. Evol. 2 , 1579–1587 (2018).
Nakamura, G., Gonçalves, L. O. & da Silva Duarte, L. Revisiting the dimensionality of biological diversity. Ecography (Cop.) 43 , 539–548 (2020).
Stevens, R. D. & Tello, J. S. On the measurement of dimensionality of biodiversity. Glob. Ecol. Biogeogr. 23 , 1115–1125 (2014).
Manning, P. et al. Simple measures of climate, soil properties and plant traits predict national-scale grassland soil carbon stocks. J. Appl. Ecol. 52 , 1188–1196 (2015).
Adler, D. & Kelly, T. vioplot: Violin plot. R package version 0.3.0 (2018).
Loreau, M. & Hector, A. Partitioning selection and complementarity in biodiversity experiments. Nature 412 , 72–76 (2001).
Allan, E. et al. Land use intensification alters ecosystem multifunctionality via loss of biodiversity and changes to functional composition. Ecol. Lett. 18 , 834–843 (2015).
Le Bagousse-Pinguet, Y. et al. Phylogenetic, functional, and taxonomic richness have both positive and negative effects on ecosystem multifunctionality. Proc. Natl Acad. Sci. USA 116 , 8419–8424 (2019).
Venail, P. et al. Species richness, but not phylogenetic diversity, influences community biomass production and temporal stability in a re-examination of 16 grassland biodiversity studies. Funct. Ecol. 29 , 615–626 (2015).
Hillebrand, H. & Matthiessen, B. Biodiversity in a complex world: consolidation and progress in functional biodiversity research. Ecol. Lett. 12 , 1405–1419 (2009).
Grace, J. B. et al. Integrative modelling reveals mechanisms linking productivity and plant species richness. Nature 529 , 390–393 (2016).
Liang, J. et al. Positive biodiversity–productivity relationship predominant in global forests. Science 354 , aaf8957 (2016).
Oehri, J., Schmid, B., Schaepman-Strub, G. & Niklaus, P. A. Biodiversity promotes primary productivity and growing season lengthening at the landscape scale. Proc. Natl Acad. Sci. USA 114 , 10160–10165 (2017).
Díaz, S. et al. Incorporating plant functional diversity effects in ecosystem service assessments. Proc. Natl Acad. Sci. USA 104 , 20684–20689 (2007).
Lavorel, S. et al. Using plant functional traits to understand the landscape distribution of multiple ecosystem services. J. Ecol. 99 , 135–147 (2011).
Schmid, B. The species richness–productivity controversy. Trends Ecol. Evol. 17 , 113–114 (2002).
Loreau, M. Biodiversity and ecosystem functioning: a mechanistic model. Proc. Natl Acad. Sci. USA 95 , 5632–5636 (1998).
Maestre, F. T. et al. Plant species richness and ecosystem multifunctionality in global drylands. Science 335 , 214–218 (2012).
van der Plas, F. et al. Jack-of-all-trades effects drive biodiversity–ecosystem multifunctionality relationships in European forests. Nat. Commun. 7 , 11109 (2016).
Socher, S. A. et al. Direct and productivity-mediated indirect effects of fertilization, mowing and grazing on grassland species richness. J. Ecol. 100 , 1391–1399 (2012).
Hobbs, R. J., Higgs, E. & Harris, J. A. Novel ecosystems: implications for conservation and restoration. Trends Ecol. Evol. 24 , 599–605 (2009).
Klaus, V. H. et al. Do biodiversity–ecosystem functioning experiments inform stakeholders how to simultaneously conserve biodiversity and increase ecosystem service provisioning in grasslands? Biol. Conserv. 245 , 108552 (2020).
Roscher, C. et al. Convergent high diversity in naturally colonized experimental grasslands is not related to increased productivity. Perspect. Plant Ecol. Evol. Syst. 20 , 32–45 (2016).
Ellenberg, H. & Leuschner, C. Vegetation Mitteleuropas mit den Alpen: In Ökologischer, Dynamischer und Historischer Sicht (UTB, 2010).
Blüthgen, N. et al. A quantitative index of land-use intensity in grasslands: integrating mowing, grazing and fertilization. Basic Appl. Ecol. 13 , 207–220 (2012).
Tilman, D., Reich, P. B. & Knops, J. M. H. Biodiversity and ecosystem stability in a decade-long grassland experiment. Nature 441 , 629–632 (2006).
Tilman, D. Community invasibility, recruitment limitation, and grassland biodiversity. Ecology 78 , 81–92 (1997).
Catford, J. A. et al. Traits linked with species invasiveness and community invasibility vary with time, stage and indicator of invasion in a long-term grassland experiment. Ecol. Lett. 22 , 593–604 (2019).
Fargione, J. et al. From selection to complementarity: shifts in the causes of biodiversity–productivity relationships in a long-term biodiversity experiment. Proc. R. Soc. B 274 , 871–876 (2007).
Londo, G. The decimal scale for releves of permanent quadrats. Vegetatio 33 , 61–64 (1976).
Roscher, C. et al. What happens to the sown species if a biodiversity experiment is not weeded? Basic Appl. Ecol. 14 , 187–198 (2013).
Kattge, J. et al. TRY—a global database of plant traits. Glob. Change Biol. 17 , 2905–2935 (2011).
Cayuela, L., Stein, A. & Oksanen, J. Taxonstand: Taxonomic standardization of plant species names. R package version 2.1 (2017).
The Plant List version 1.1 (2013); http://www.theplantlist.org/
Qian, H. & Jin, Y. An updated megaphylogeny of plants, a tool for generating plant phylogenies and an analysis of phylogenetic community structure. J. Plant Ecol. 9 , 233–239 (2016).
Martins, W. S., Carmo, W. C., Longo, H. J., Rosa, T. C. & Rangel, T. F. SUNPLIN: simulation with uncertainty for phylogenetic investigations. BMC Bioinform. 14 , 324 (2013).
Rangel, T. F. et al. Phylogenetic uncertainty revisited: implications for ecological analyses. Evolution 69 , 1301–1312 (2015).
Cornelissen, J. H. C. et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust. J. Bot. 51 , 335–380 (2003).
Goolsby, E. W., Bruggeman, J. & Ane, C. Rphylopars: Phylogenetic comparative tools for missing data and within-species variation. R package version 0.2.9 (2016).
Penone, C. et al. Imputation of missing data in life-history trait datasets: which approach performs the best? Methods Ecol. Evol. 5 , 961–970 (2014).
Oksanen, J. et al. Vegan: Community ecology package. R package version 2.3-4 (2016).
Hill, M. Diversity and evenness: a unifying notation and its consequences. Ecology 54 , 427–432 (1973).
Smith, B. & Wilson, J. B. A consumer’s guide to evenness indices. Oikos 76 , 70–82 (1996).
Magurran, A. Measuring Biological Diversity (Blackwell, 2004).
Morris, E. K. et al. Choosing and using diversity indices: insights for ecological applications from the German biodiversity exploratories. Ecol. Evol. 4 , 3514–3524 (2014).
Tucker, C. M. et al. A guide to phylogenetic metrics for conservation, community ecology and macroecology. Biol. Rev. 92 , 698–715 (2017).
Kembel, S. W. et al. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 26 , 1463–1464 (2010).
Villéger, S., Mason, N. W. H. & Mouillot, D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology 89 , 2290–2301 (2008).
Laliberte, E. & Legendre, P. A distance-based framework for measuring functional diversity from multiple traits. Ecology 91 , 299–305 (2010).
Mouchet, M. A., Villéger, S., Mason, N. W. H. & Mouillot, D. Functional diversity measures: an overview of their redundancy and their ability to discriminate community assembly rules. Funct. Ecol. 24 , 867–876 (2010).
Laliberté, E., Legendre, P. & Shipley, B. FD: Measuring functional diversity from multiple traits, and other tools for functional ecology. R package version 1.0-12 (2014).
R: A Language and Environment for Statistical Computing v.3.4.2 (R Core Team, 2019); https://doi.org/10.1007/978-3-540-74686-7
Dormann, C. F. et al. Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography (Cop.) 36 , 27–46 (2013).
Zuur, A. F., Ieno, E. N. & Elphick, C. S. A protocol for data exploration to avoid common statistical problems. Methods Ecol. Evol. 1 , 3–14 (2010).
Jochum, M. et al. R-code and aggregated data from: The results of biodiversity-ecosystem functioning experiments are realistic. iDiv Data Repository https://doi.org/10.25829/idiv.1869-11-3082 (2020).
Fox, J. & Weisberg, S. An R Companion to Applied Regression (SAGE, 2011).
Pebesma, E. & Bivand, R. Classes and methods for spatial data in R. R News 5 , 9–13 (2005).
Bivand, R. S., Pebesma, E. & Gomez-Rubio, V. Applied Spatial Data Analysis with R (Springer, 2013).
Habel, K., Grasman, R., Gramacy, R. B., Stahel, A. & Sterratt, D. C. geometry: Mesh generation and surface tessellation. R package version 0.4.1 (2019).
Blonder, B. & Harris, D. hypervolume: High dimensional geometry and set operations using kernel density estimation, support vector machines, and convex hulls. R package version 2.0.11 (2018).
Meyer, S. T. et al. Effects of biodiversity strengthen over time as ecosystem functioning declines at low and increases at high biodiversity. Ecosphere 7 , e01619 (2016).
Brownrigg, R. mapdata: Extra map databases. R package version 2.3.0 (2018).
Download references
Acknowledgements
We thank the establishers, maintainers, coordinators, technical and research staff, and data owners of all involved projects, as well as the TRY initiative. We thank S. Soliveres and E. Allan for discussion; S. Thiel, G. Luo, D. Bahauddin and F. Schneider for help with data extraction and handling; and R. Junker and B. Blonder for assistance with the calculation of multidimensional hypervolumes. This study was funded through Jena Experiment SP 7 (Swiss National Science Foundation grant no. 310030E-166017/1). Further support came from the German Centre for Integrative Biodiversity Research (iDiv) Halle-Jena-Leipzig, funded by the German Research Foundation (grant no. FZT 118). The Jena Experiment was funded by the Deutsche Forschungsgemeinschaft (grant nos FOR 456 and FOR 1451) with additional support from Friedrich Schiller University Jena, the Max Planck Institute for Biogeochemistry in Jena and the Swiss National Science Foundation. All Cedar Creek studies are funded by the US National Science Foundation’s Long-Term Ecological Research (LTER) programme (grant no. DEB-1234162). F.I. acknowledges funding from the LTER Network Communications Office (grant no. DEB-1545288). We thank the iDiv Data Repository for hosting our R code and aggregated datasets and for performing the related quality checks.
Author information
Authors and affiliations.
Institute of Plant Sciences, University of Bern, Bern, Switzerland
Malte Jochum, Markus Fischer, Steffen Boch, Rafael Molina-Venegas, Caterina Penone, Daniel Prati, Abiel Rindisbacher & Deborah Schäfer
German Centre for Integrative Biodiversity Research (iDiv) Halle-Jena-Leipzig, Leipzig, Germany
Malte Jochum, Christiane Roscher, Nico Eisenhauer, Jens Kattge, Anja Vogel & Alexandra Weigelt
Institute of Biology, Leipzig University, Leipzig, Germany
Malte Jochum, Nico Eisenhauer & Anja Vogel
Department of Ecology, Evolution and Behavior, University of Minnesota, Saint Paul, MN, USA
Forest Isbell, Jeannine Cavender-Bares & David Tilman
Department of Physiological Diversity, UFZ, Helmholtz Centre for Environmental Research, Leipzig, Germany
Christiane Roscher
Systematic Botany and Functional Biodiversity, Institute of Biology, Leipzig University, Leipzig, Germany
Fons van der Plas & Alexandra Weigelt
WSL Swiss Federal Research Institute, Birmensdorf, Switzerland
Steffen Boch
Max Planck Institute for Biogeochemistry, Jena, Germany
Gerhard Boenisch, Gerd Gleixner, Jens Kattge & Markus Lange
Institute of Agricultural Sciences, ETH Zürich, Zürich, Switzerland
Nina Buchmann & Valentin H. Klaus
Department of Geography, King’s College London, London, UK
Jane A. Catford
Institute of Ecology and Evolution, University of Jena, Jena, Germany
Anne Ebeling & Anja Vogel
Institute of Landscape Ecology, University of Münster, Münster, Germany
Norbert Hölzel
Institute of Landscape Ecology and Resources Management, Giessen University, Giessen, Germany
Till Kleinebecker
Senckenberg Biodiversity and Climate Research Centre (SBiK-F), Frankfurt, Germany
Gaëtane Le Provost & Peter Manning
Terrestrial Ecology Research Group, School of Life Sciences Weihenstephan, Technical University of Munich, Freising, Germany
Sebastian T. Meyer & Wolfgang W. Weisser
GloCEE–Global Change Ecology & Evolution group, Department of Life Sciences, University of Alcalá, Madrid, Spain
Rafael Molina-Venegas
Plant Ecology and Nature Conservation Group, Wageningen University and Research, Wageningen, The Netherlands
Liesje Mommer
Geoecology, Geoscience Department, University of Tübingen, Tübingen, Germany
Yvonne Oelmann
Department of Forest Resources, University of Minnesota, Saint Paul, MN, USA
Peter B. Reich
Hawkesbury Institute for the Environment, Western Sydney University, Penrith South DC, New South Wales, Australia
JF Blumenbach Institute of Zoology and Anthropology, University of Göttingen, Göttingen, Germany
Stefan Scheu
Centre for Biodiversity and Sustainable Land Use, University of Göttingen, Göttingen, Germany
Department of Geography, University of Zürich, Zürich, Switzerland
Bernhard Schmid
Institute of Ecology, College of Urban and Environmental Sciences, Peking University, Beijing, China
Bren School of Environmental Science and Management, University of California, Santa Barbara, CA, USA
David Tilman
Agroecology, Department of Crop Sciences, University of Göttingen, Göttingen, Germany
Teja Tscharntke
Department of Evolutionary Biology and Environmental Studies, University of Zürich, Zürich, Switzerland
Cameron Wagg
Institute of Geography and Geoecology, Karlsruhe Institute of Technology (KIT), Karlsruhe, Germany
Wolfgang Wilcke
You can also search for this author in PubMed Google Scholar
Contributions
M.J., P.M., M.F. and F.v.d.P. conceived and designed the study. M.J., M.F., F.I., C.R., S.B., G.B., N.B., J.A.C., J.C.-B., A.E., N.E., G.G., N.H., J.K., V.H.K., T.K., M.L., G.L.P., S.T.M., L.M., Y.O., D.P., P.B.R., D.S., S.S., B.S., D.T., T.T., A.V., C.W., A.W., W.W.W., W.W. and P.M. contributed data. M.J. developed the analytical framework and analysed the data. R.M.-V. constructed the phylogenetic hypothesis trees. M.J. and P.M. wrote the manuscript. M.J., M.F., F.I., C.R., F.v.d.P., S.B., G.B., N.B., J.A.C., J.C.-B., A.E., N.E., G.G., N.H., J.K., V.H.K., T.K., M.L., G.L.P., S.T.M., R.M.-V., L.M., Y.O., C.P., D.P., P.B.R., A.R., D.S., S.S., B.S., D.T., T.T., A.V., C.W., A.W., W.W.W., W.W. and P.M. contributed to the discussion of the results and writing of the manuscript.
Corresponding author
Correspondence to Malte Jochum .
Ethics declarations
Competing interests.
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended data fig. 1 list of german and us datasets for vegetation and ecosystem function data..
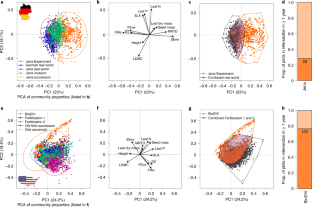
Lists country, project name, project code used in this paper, main reference, number of plots we used, years we have vegetation data for, functions we used including years. Most of the raw data is openly available in various online repositories: Jena Experiment ( http://jenaexperiment.uni-jena.de/index.php/data/ ), Biodiversity Exploratories ( https://www.bexis.uni-jena.de/Login/Account.aspx ), Cedar Creek ( https://www.cedarcreek.umn.edu/research/data ). Data from the Saale grasslands (Jena real world) were provided by Christiane Roscher and are currently not openly available. Aggregated datasets used for this study are now available online 101 .
Extended Data Fig. 2 Temporal movement of Jena invasion communities into the real-world realm.
Based on the PCA in Fig. 1a . Different shades of purple show Jena invasion communities across the years from 2003-2009. Orange and gray ellipses show 95% confidence intervals for Jena Experiment and combined real-world plots (but their communities are not plotted here), respectively. Note that while the points in different panels are from single years, the ellipses are fixed to the across-year comparison in Fig. 1a . The last panel shows the PCA factor loadings for the 12 vif-selected community properties (arrows scaled to improve visibility - “const=25” in R vegan “biplot” function 87 ). Within six years of succession, the plant communities of Jena invasion plots fully “moved” into the core of the community property space defined by the combined real-world plots (German real world and Jena real world, respectively).
Extended Data Fig. 3 Violin plots of all 21 community properties of German data.
Experimental (E, Jena Experiment, strong orange, 82 plots), unrealistic experimental (unreal., intermediate orange, 59 plots), selected realistic experimental (real., weak orange, 23 plots) and combined real-world plots (German real world, Jena real world, gray, 164 plots), all averaged across years per plot. Combination of boxplot and rotated kernel density plot (R package “vioplot” 51 ). Realistic plots were calculated based on the 12 vif-selected community properties and the convex hull volume method. Units: leaf area (mm²), leaf dry mass (mg), leaf dry matter content (LDMC, g/g), leaf nitrogen concentration (leaf N, mg/g), leaf phosphorus concentration (leaf P, mg/g), plant height (m), specific leaf area (SLA, mm²/mg) and seed mass (dry mass in mg). Other community properties are dimensionless.
Extended Data Fig. 4 Violin plots of all 21 community properties of US data.
Experimental (E, BioDIV, strong orange, 159 plots), unrealistic experimental (unreal., intermediate orange, 37 plots), selected realistic experimental (real., weak orange, 122 plots) and combined real-world plots (Fertilization 1 & 2, gray, 369 plots), all averaged across years per plot. Combination of boxplot and rotated kernel density plot (R package “vioplot” 51 ). Realistic plots were calculated based on the 12 vif-selected community properties and the convex hull volume method. Units: leaf area (mm²), leaf dry mass (mg), leaf dry matter content (LDMC, g/g), leaf nitrogen concentration (leaf N, mg/g), leaf phosphorus concentration (leaf P, mg/g), plant height (m), specific leaf area (SLA, mm²/mg) and seed mass (dry mass in mg). Other community properties are dimensionless.
Extended Data Fig. 5 Model parameters for BEF relationships presented in Fig. 2 .
Values are presented for unconstrained (u) and constrained (c) models of Jena (J) and BioDIV BEF relationships. Constraining was done using the 12 vif-selected community properties and the convex hull method. Sample size (n), slope estimates (slop), lower (low) and upper (upp) 95% confidence intervals, p-values (p) and adjusted R 2 values (R2). All values are rounded to two decimal places.
Extended Data Fig. 6 Variance explained by 12 PCA axes (12 vif-selected community properties).
Percentage of total variance explained by each of the 12 PCA axes (PC’s, see Fig. 1 ) for each region (GER = Germany and US = USA). Rounded to two decimal places.
Extended Data Fig. 7 PCA scores for 12 vif-selected community properties of PCA’s in Fig. 1 .
Scores have been produced using the scores() command of the “vegan” package 87 in R and have been rounded to two decimal places.
Supplementary information
Supplementary information.
Supplementary Figs. 1–10, Tables 1–17, Supplementary Information on Sensitivity Analyses 1 and 2, and methods.
Reporting Summary
Rights and permissions.
Reprints and permissions
About this article
Cite this article.
Jochum, M., Fischer, M., Isbell, F. et al. The results of biodiversity–ecosystem functioning experiments are realistic. Nat Ecol Evol 4 , 1485–1494 (2020). https://doi.org/10.1038/s41559-020-1280-9
Download citation
Received : 31 July 2019
Accepted : 17 July 2020
Published : 24 August 2020
Issue Date : November 2020
DOI : https://doi.org/10.1038/s41559-020-1280-9
Share this article
Anyone you share the following link with will be able to read this content:
Sorry, a shareable link is not currently available for this article.
Provided by the Springer Nature SharedIt content-sharing initiative
This article is cited by
Tree and shrub richness modifies subtropical tree productivity by regulating the diversity and community composition of soil bacteria and archaea.
Microbiome (2023)
Reply: Functional similarity is more appropriate than functional redundancy
- Nico Eisenhauer
- Matthias C. Rillig
npj Biodiversity (2023)
Relationships between species richness and biomass production are context dependent in grasslands differing in land-use and seed addition
- Karl Andraczek
- Alexandra Weigelt
- Fons van der Plas
Scientific Reports (2023)
Local-scale soil nematode diversity in a subtropical forest depends on the phylogenetic and functional diversity of neighbor trees
- Xiaofang Du
Plant and Soil (2023)
Thresholds and tipping points are tempting but not necessarily suitable concepts to address anthropogenic biodiversity change—an intervention
- Helmut Hillebrand
- Lucie Kuczynski
- Jan-Claas Dajka
Marine Biodiversity (2023)
Quick links
- Explore articles by subject
- Guide to authors
- Editorial policies
Sign up for the Nature Briefing newsletter — what matters in science, free to your inbox daily.

Click through the PLOS taxonomy to find articles in your field.
For more information about PLOS Subject Areas, click here .
Loading metrics
Open Access
Peer-reviewed
Research Article
Scientific research on animal biodiversity is systematically biased towards vertebrates and temperate regions
Roles Data curation, Formal analysis, Investigation, Methodology, Project administration, Resources, Software, Validation, Visualization, Writing – original draft, Writing – review & editing
* E-mail: [email protected]
Affiliation Insect Ecology Group, University Museum of Zoology, Cambridge, Downing Street, Cambridge, United Kingdom
Roles Conceptualization, Writing – review & editing
Affiliation Biological Sciences, Institute for Life Sciences, University of Southampton, Southampton, United Kingdom
Roles Conceptualization, Funding acquisition, Methodology, Supervision, Writing – review & editing
- Mark A. Titley,
- Jake L. Snaddon,
- Edgar C. Turner
- Published: December 14, 2017
- https://doi.org/10.1371/journal.pone.0189577
- Reader Comments
Over the last 25 years, research on biodiversity has expanded dramatically, fuelled by increasing threats to the natural world. However, the number of published studies is heavily weighted towards certain taxa, perhaps influencing conservation awareness of and funding for less-popular groups. Few studies have systematically quantified these biases, although information on this topic is important for informing future research and conservation priorities. We investigated: i) which animal taxa are being studied; ii) if any taxonomic biases are the same in temperate and tropical regions; iii) whether the taxon studied is named in the title of papers on biodiversity, perhaps reflecting a perception of what biodiversity is; iv) the geographical distribution of biodiversity research, compared with the distribution of biodiversity and threatened species; and v) the geographical distribution of authors’ countries of origin. To do this, we used the search engine Web of Science to systematically sample a subset of the published literature with ‘biodiversity’ in the title. In total 526 research papers were screened—5% of all papers in Web of Science with biodiversity in the title. For each paper, details on taxonomic group, title phrasing, number of citations, study location, and author locations were recorded. Compared to the proportions of described species, we identified a considerable taxonomic weighting towards vertebrates and an under-representation of invertebrates (particularly arachnids and insects) in the published literature. This discrepancy is more pronounced in highly cited papers, and in tropical regions, with only 43% of biodiversity research in the tropics including invertebrates. Furthermore, while papers on vertebrate taxa typically did not specify the taxonomic group in the title, the converse was true for invertebrate papers. Biodiversity research is also biased geographically: studies are more frequently carried out in developed countries with larger economies, and for a given level of species or threatened species, tropical countries were understudied relative to temperate countries. Finally, biodiversity research is disproportionately authored by researchers from wealthier countries, with studies less likely to be carried out by scientists in lower-GDP nations. Our results highlight the need for a more systematic and directed evaluation of biodiversity studies, perhaps informing more targeted research towards those areas and taxa most depauperate in research. Only by doing so can we ensure that biodiversity research yields results that are relevant and applicable to all regions and that the information necessary for the conservation of threatened species is available to conservation practitioners.
Citation: Titley MA, Snaddon JL, Turner EC (2017) Scientific research on animal biodiversity is systematically biased towards vertebrates and temperate regions. PLoS ONE 12(12): e0189577. https://doi.org/10.1371/journal.pone.0189577
Editor: Bernd Schierwater, Tierarztliche Hochschule Hannover, GERMANY
Received: February 5, 2017; Accepted: November 29, 2017; Published: December 14, 2017
Copyright: © 2017 Titley et al. This is an open access article distributed under the terms of the Creative Commons Attribution License , which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Data Availability: All relevant data are within the paper and its Supporting Information files.
Funding: The authors received no specific funding for this work.
Competing interests: The authors have declared that no competing interests exist.
Introduction
Since 1988, when the word was first used in a publication [ 1 ], the idea of ‘biodiversity’ has become integrated into both popular and scientific culture. The word produces more than 50 million hits on Google [ 2 ] and almost 90,000 in the scientific search engine and database Web of Science at the time of writing [ 3 ]. Moreover, systematic quantification of the number of papers studying biodiversity shows a marked increase over the last two decades ( Fig 1 ).
- PPT PowerPoint slide
- PNG larger image
- TIFF original image
A search for the word ‘biodiversity’ in Web of Science by year reveals the increase in biodiversity research over time (search date: 10 th February 2016).
https://doi.org/10.1371/journal.pone.0189577.g001
Biodiversity was formally defined at the 1992 United Nations Convention on Biological Diversity as ‘the variability among living organisms from all sources including, inter alia, terrestrial, marine and other aquatic ecosystems and the ecological complexes of which they are part; this includes diversity within species, between species and of ecosystems’[ 4 ]. The most commonly used meaning is diversity at the species level, although despite being an intuitive concept, in practice definitions of what constitutes a species, and estimates of Earth’s species richness, remain uncertain and variable. Estimates for global species richness typically fall in the range of 3 million to 100 million species [ 5 ] although a working figure between 5 and 15 million is often suggested [ 6 ].
Contrary to this uncertainly, it is well established that diversity is not evenly distributed amongst taxa. Arthropods, and especially insects, account for most known eukaryote species: of the 1.2–2 million described species, approximately 925,000 are insects [ 7 , 8 ]. However, it has become clear that public perceptions of biodiversity do not reflect this invertebrate-dominated reality. In the UK, children asked to draw their ‘ideal rainforest’ over-represented mammals, reptiles and birds, and under-represented insects and annelids [ 9 ]. Such taxonomic chauvinism is by no means restricted to children, nor is it restricted to non-academics: 31% of papers published in 2001 in three prominent conservation journals focussed on birds and mammals [ 10 ]. Although this focus on larger species is understandable, owing to their greater apparency and potentially greater importance for ecosystem processes and vulnerability to environmental change [ 11 , 12 ], it does mean that invertebrate conservation issues and extinctions may go unreported or unacknowledged. This could hamper an overarching understanding of the state of the natural environment. For example, only 70 modern insect extinctions have been documented, despite thousands being estimated to have occurred [ 13 ].
Several previous studies have examined these taxonomic biases in journal articles. A survey of papers on vertebrates from nine high-impact journals reported a bias towards mammals and birds [ 14 ]. Furthermore, mammal and bird studies had more ‘narrowly framed’ introductions and mentioned the study organisms sooner than in studies on fishes, reptiles or amphibians. In a review of fifteen years of research from two leading conservation research journals ( Biological Conservation and Conservation Biology ), an over-representation of vertebrates and under-representation of invertebrates was revealed [ 15 ]. Within vertebrates, birds and mammals were over-represented, while other taxa were under-represented. A similar study analysed the research in three prominent conservation journals [ 10 ], finding once again a weighting in favour of vertebrates, as well as towards pristine landscapes and single species, rather than communities. Another study focussed on the research output of four ecological journals ( Journal of Animal Ecology , Journal of Applied Ecology , Oecologia , Ecology ) for the years 2006 and 2007 [ 16 ], and again highlighted the tendency to ignore invertebrates, in particular insects, in high-impact journals. Also reported was a preference in British Research Council NERC funding towards vertebrate ecologists (38%) compared with entomologists (13%).
Thus, the topic of taxonomic seems well studied, although these four papers all used a similar approach, focussing on the research output of a few selected journals. In the present article, we take a different, more wide-ranging approach, sampling across the published literature for papers whose title contains the word biodiversity. We therefore do not discriminate by journal (hence nor by impact factor), aiming to obtain a more holistic and longer-term view of taxonomic biases in global biodiversity research. In addition, we chose to investigate geographical biases, to assess whether biodiversity research is skewed towards certain regions and whether taxonomic biases are stronger in certain parts of the world.
Specifically, we first investigate whether reported taxonomic biases (towards vertebrates, and towards birds and mammals especially) pervade papers on biodiversity and whether this weighting has changed over time. Secondly, we investigate whether any bias differs between temperate and tropical regions. Thirdly, we investigate how the titles of papers on biodiversity are phrased. In particular, whether papers studying biodiversity differ in how likely they are to specify the study taxon in the title compared between papers on invertebrate and vertebrate biodiversity. This may reflect and promote a common (if subconscious) perception of which taxa represent biodiversity. Fourthly, we investigate the global distribution of biodiversity research, compared to the actual distribution of biodiversity, to assess how well research effort reflects biodiversity. We also compare it to the distribution of IUCN Red-Listed species and GDP, to assess how research effort reflects conservation priorities and wealth. Finally, we investigate the authors’ countries of origin relative to the study location, to assess whether there is a mismatch between the distribution of research on biodiversity and biodiversity researchers by country.
Materials & methods
Sample selection.
The scientific citation-indexing platform ‘Web of Science’ was used to sample research papers from the period 1995–2015, following a strict and repeatable search protocol. To be eligible for inclusion, papers’ title must have contained the word ‘biodiversity’, and also had to be a primary research article, in order to exclude review papers and other publication types such as books (which might have led to double-counting of studies). For each year, we then randomly selected 5% of all eligible articles using the random number generator www.random.org [ 17 ]. Five percent was an arbitrary figure that produced a sample size of 526 publications, which was quantifiable within the time frame of this project. This method may be cruder and return more irrelevant results than the careful examination of selected journals, but enabled us to easily generate a large sample size, and sample across a broad range of journals and disciplines over many years to obtain a comprehensive selection of biodiversity research. In this study we chose to focus on biases in animal biodiversity research, although we acknowledge that biases may also exist and be important across other taxonomic groups.
Data collection
For each of the 526 papers in our sample, we recorded the taxon/taxa studied; the climate zone (temperate or tropical) in which the study took place; whether or not the taxonomic group was specified in the title; the country in which the study took place; the country of origin of the paper’s authors; and the number of times that paper had been cited as recorded in Web of Science at the time of searching. Vertebrate studies were classified into one or more of five major vertebrate groups (Mammals, Birds, Reptiles, Amphibians and Fishes). Correspondingly, five major invertebrate groups were chosen because of their high species richness and because they are relatively well studied (Insects, Arachnids, Nematodes, Annelids, and Molluscs). Studies on invertebrates that could not be classified into these five groups were recorded as ‘Other invertebrates’. When recording the climate zone, we considered any studies taking place between the Tropics of Cancer and Capricorn (23.5°N and S respectively) as ‘tropical’. Since only six polar studies existed in the sample, there were not enough to include these as a separate climate zone. We therefore considered all studies taking place at latitudes higher than the tropics to be ‘temperate’. By this classification, studies in polar regions are also classified as temperate. For each author, their country of origin was recorded as the country of their affiliated institution. If a paper had multiple authors from different countries, multiple countries were recorded for the authors’ country of origin.
Data analysis
Statistical analyses were performed using R (version 3.0.2) [ 18 ]. To analyse the top 25% most-cited papers separately, the average number of citations per year was calculated (total citations to date divided by the time since publication). Chi-square tests were used to test for differences between temperate and tropical regions, and whether taxa were specified or not in the title. Wilcoxon rank-sum tests were used to test for differences between vertebrate and invertebrate residuals when comparing taxa for the proportion of studies versus proportion of described species as listed on the International Union for the Conservation of Nature (IUCN) database. Generalised linear models were used to test whether the number of biodiversity studies or authors in a country was related to Gross Domestic Product (GDP)–data from World Bank : World Development Indicators 2014 . Maps were created using QGIS (version 2.12.1) to visualise differences in research effort across countries worldwide. In particular, we mapped the number of biodiversity publications per 1000km 2 on vertebrates and invertebrates for each country, to visualise biases in research effort. We also mapped the number of authors relative to each country’s human population. By dividing the number of threatened species (data from IUCN [ 19 ]) by the number of biodiversity papers for each country, we also visualised countries that could be considered priorities for research (high numbers of threatened species relative to biodiversity research effort). Finally, analysis of covariance (ANCOVA) was used to test whether tropical and temperate regions differed in research effort for a given level of species or threatened species.
Taxonomic biases
Approximately half of the papers sampled studied vertebrates, and half studied invertebrates ( Fig 2 ). However, this is far from the true proportions of described species, where over 95% of species are invertebrates (see right-hand column of Fig 2 ). Furthermore, this focus on vertebrates has been roughly consistent over the last 20 years. Given their true species richness, vertebrates were significantly over-represented compared to invertebrates in the published literature (Wilcoxon rank-sum test, W = 24, N = 10, P<0.05) ( Fig 3 ). Invertebrate taxa were either slightly over-represented (annelids, molluscs, nematodes and ‘other invertebrates’) or under-represented (insects and arachnids). In addition, the taxonomic bias was greater in highly cited papers. Of the top 25% most cited papers in the sample, only 47% included invertebrates, compared with 57% of the entire sample.
The proportion of different taxonomic groups in the sample of papers with ‘biodiversity’ in the title is shown for 4 five-year periods since 1996. For comparison, the right-hand column illustrates the ‘true’ proportions of described species that each group makes up (data from IUCN [ 20 ]) Vertebrate and invertebrate taxa are separated by a grey line.
https://doi.org/10.1371/journal.pone.0189577.g002
The proportion of studies on each taxonomic group is plotted against the ‘actual’ proportion of described species [ 20 ] found in that taxon. Values were log transformed for clarity. The 1:1 line is shown (dotted); over-represented groups are found above the line while under-represented groups are below it. Vertebrate groups are shown in red and invertebrate groups are shown in blue.
https://doi.org/10.1371/journal.pone.0189577.g003
Comparing tropical and temperate regions
In terms of the proportion of studies, the bias towards vertebrates was greater in tropical regions than temperate regions (Chi-square test, X 2 = 30.65, N = 672, P<0.001) ( Fig 4 ). In tropical countries, 43% of studies included invertebrates, compared to 63% in temperate countries. General patterns of taxonomic over- or under-representation were similar in tropical and temperate regions, although arachnids were particularly under-represented in the tropics, and molluscs were under-represented in the tropics despite being over-represented in temperate studies.
The bias towards vertebrates is greater in tropical regions than temperate regions. The proportions of described species in different groups are shown in the right-hand column for comparison.
https://doi.org/10.1371/journal.pone.0189577.g004
Differences in title phrasing
The proportion of papers for which a taxonomic group was specified in the title differed between vertebrates and invertebrates (Chi-square test, X 2 = 103.45, N = 714, P<0.0001) ( Fig 5 ). Specifically, most papers that studied vertebrates did not specify the study taxon/taxa in the title, and instead referred to ‘biodiversity’ more generally. In contrast, the titles of studies on invertebrates usually specified which taxa were being studied. An exception to this pattern was studies on fishes, where the majority of studies specified the taxon in the title.
The majority of studies on vertebrates (with the exception of studies on fishes) do not mention the study taxon in the title. Conversely, for papers on invertebrates, the taxa being studied were specified more often than not.
https://doi.org/10.1371/journal.pone.0189577.g005
Geographic biases
Biodiversity research was more commonly carried out in developed countries with larger economies, for both vertebrate and invertebrate studies ( Fig 6 ). The United States of America had the highest number of studies of any country in the sample, but the density of biodiversity research appears to be generally highest in Western Europe. Most tropical areas had fewer studies and very little research was based in African countries. The number of biodiversity studies was positively related to countries’ nominal GDP (Poisson regression, z = 28.62, N = 232, P<0.0001) ( Fig 7 ).
The number of papers with ‘biodiversity’ in the title per 1000km 2 is shown, for a) papers that study vertebrates and b) papers that study invertebrates. Darker colours represent a higher density of studies.
https://doi.org/10.1371/journal.pone.0189577.g006
Nominal GDP in US$ is plotted against the number of biodiversity studies sampled from each country, revealing a positive relationship. The top ten countries for number of papers are labelled. Many countries with low GDP had no biodiversity papers identified from this sample.
https://doi.org/10.1371/journal.pone.0189577.g007
Certain counties had a higher number of threatened species relative to the biodiversity research effort (given by dividing the number of IUCN listed threatened species [ 19 ] by the number of research publications on biodiversity ( Fig 8 ). In particular, northern South America, Africa and SE Asia had a low relative number of publications. Note that large areas of Africa lacked any studies at all in our sample. We recorded a generally a positive relationship between the number of publications and the number of threatened and number of species recorded in the IUCN database [ 19 , 20 ] per country. However, for a given level of species or threatened species, tropical regions were understudied compared to temperate regions; interactions were significant between climate region and number of threatened species (F 3,227 = 36.06, p<0.0001) ( Fig 9A ) and between climate region and number of species (F 3 , 227 = 48.28, p<0.0001) ( Fig 9B ).
Dividing the number of animal species threatened with extinction [ 19 ] by the number of biodiversity studies reveals regions that are understudied given their number of threatened species. Countries in northern South America, Africa and SE Asia stand out as being relatively understudied; much of central Africa lacked studies altogether in this sample. Darker colours represent a higher number of listed threatened species per study.
https://doi.org/10.1371/journal.pone.0189577.g008
Scatterplots comparing the number of biodiversity papers against the number of threatened animal species (a) and species richness (b) listed in IUCN databases [ 19 , 20 ] per country. Temperate countries tend to have more biodiversity research than tropical countries for a given number of threatened species or a given species richness.
https://doi.org/10.1371/journal.pone.0189577.g009
As with the distribution of biodiversity research, the distribution of authors was heavily biased towards developed countries, particularly Western Europe ( Fig 10 ). Many countries in Africa, central Asia and South America lacked any authors on the papers in the sample; this is particularly true when looking at lead authors only ( Fig 10B ). The number of authors from a country was strongly related to wealth of that country as approximated by nominal GDP (Poisson regression, z = 69.91, N = 232, P < 0.0001). Furthermore, the GDP of authors’ countries of origin (median 2,066,902 million US$) was significantly higher than the GDP of study locations (median 1,453,770 million US$) (Wilcoxon rank-sum test, N = 513, W = 89086, P < 0.0001).
The number of authors (a) and lead authors (b) from each country relative to the country’s population. Many countries in Africa, central Asia and South America lacked authors on the papers in the sample.
https://doi.org/10.1371/journal.pone.0189577.g010
The taxonomic bias
These results clearly demonstrate more charismatic animal groups are over-represented in biodiversity research and have been since biodiversity first emerged as a research field. Mammals, which make up around 0.4% of known animal species [ 20 ], were studied in approximately 12% of papers with biodiversity in the title. The equivalent numbers for birds are 0.7% and 13%. In contrast, insects make up at least 70% of animal species [ 20 ] yet were studied in less than a quarter (23%) of papers. This result corroborates earlier findings [ 10 , 14 – 16 ], and extends the phenomenon to all biodiversity research rather than just that of selected journals. Due to the high proportion of species remaining to be described, particularly among the invertebrates, this figure is likely to be conservative. These results have implications for awareness of the natural world in the scientific community, particularly as this taxonomic bias was greater in the top quartile of most-cited papers, suggesting that the research with the highest impact and largest influence is even less representative of the real world in this regard.
The taxonomic bias was greater in tropical regions, where vertebrates were studied in more than half of papers, despite vertebrates comprising less than 5% of animal species. As tropical countries contain a higher total species number and are therefore likely to have a much higher proportion of undescribed species [ 5 ], particularly smaller taxa, this under-representation is likely to be even more marked in reality. Ensuring adequate research coverage across taxa in tropical regions has important conservation implications. Most species are found in the tropics [ 21 ] and tropical regions encompass many of the world’s conservation priority hotspots [ 22 ], but are currently experiencing habitat loss faster than any other region [ 23 ].
Not all invertebrate taxa were underrepresented however; in fact, four out of the six invertebrate groups were somewhat over-represented in scientific research. The overall lack of invertebrate studies is, more precisely, a dearth of global insect and arachnid research and tropical mollusc research. The fact that insects and arachnids were the least well represented groups in this study does not mean they are the least represented of all taxa, since there will be other poorly studied invertebrate groups included within the other invertebrates category, or within these groups at a finer taxonomic scale. However, since arachnids and insects are so speciose, the deficiency of research in these groups is perhaps most significant to understanding global biodiversity. Another key finding relating to taxonomic bias is that studies on vertebrates typically did not specify the taxon in the title, referring to ‘biodiversity’ more generally. This was not the case for invertebrate research, for which the study taxa were usually specified. This could reflect a general perception that vertebrates alone are sufficient to represent biodiversity.
This unequal coverage of research across taxa may have a complex combination of causes. Researchers themselves may find studying charismatic vertebrates more appealing. Alternatively, it could represent the increased challenges of working with more diverse taxa, particularly in terms species identification. This is despite studies showing that certain insect groups are informative indicators of biodiversity and cost effective taxa to sample [ 24 , 25 ]. General perceptions of biodiversity may also be influenced by journal editors publishing a disproportionate number of articles on vertebrates (consciously or subconsciously), because such articles may be more likely to gain traction within a scientific community that is already vertebrate-biased (especially if journals are under pressure to maintain a high impact factor driven by citations). Vertebrate-biased research may also appeal to the media who are catering for a vertebrate-preferring public audience [ 9 ]. The taxonomic bias could also be the product of funding bodies, which may preferentially award research grants for vertebrate studies if these are perceived to be more important, interesting or relevant to conservation and policy priorities. A few or all of these hypotheses may play a role in producing the biases reported in this study.
Taxonomic bias is not necessarily bad. A bias towards charismatic vertebrate taxa may be advantageous where such taxa have a disproportionately large role in ecosystem functioning (keystone species), in generating funds and support for conservation (flagship species), or when their protection also ensures the protection of much of their ecosystem (umbrella species) [ 26 , 27 ]. In addition, certain taxa may be used as surrogates for other harder-to-study groups [ 28 , 29 ], which may have a similar geographic distribution or show a similar response to disturbance. However, notwithstanding doubt over the prevalence of keystone species and the reliability of taxonomic surrogates [ 30 , 31 ], it is unlikely that the taxonomic bias we have observed has arisen as a result of deliberate decisions to select these taxa as indicators of other lesser-known animal groups.
In using the proportion of described species as a reference for many of our analyses, we implicitly make the assumption that all species are equal. However, clearly this is not the case in terms of ecosystem function or conservation priority. It would be interesting to investigate whether the proportion of research done on different taxonomic groups better reflects the distribution of ecological importance or conservation value among taxa (rather than the proportion of described species), but it remains a challenge to identify meaningful measures for these that are comparable across taxa and globally applicable [ 32 ].
The geographic bias
The distribution of biodiversity research and its authors’ countries of origin resemble the distribution of GDP, rather than that of actual biodiversity or numbers of threatened species. The distribution of research is skewed towards developed countries and particularly Western Europe. Furthermore, even when studies are carried out in lower GDP-countries, the authors tend to be based at institutions in wealthier nations. Tropical countries tend to have fewer biodiversity studies despite being where more biodiversity is found and where biodiversity is most threatened. Tropical regions were also where the taxonomic bias was greatest. Taken together, these findings have important implications for biodiversity conservation: the same areas that are most threatened and most diverse are the least studied [ 23 ] and where scientists research is most skewed towards less-speciose groups. Therefore, we are likely to continue to undervalue these under-studied groups, especially in parts of the world where they are most threatened, and perhaps allocate less funding to their protection. Moreover, given that conservation efforts will be more likely to succeed when we better understand the target organisms, there is a real possibility that we may be ill equipped to protect the majority of animal biodiversity. Research gaps may mean we are less likely to identify threatened invertebrates and notice their disappearance, and we may be less likely to identify underlying threats and their drivers. Furthermore, without a good understanding of invertebrate biodiversity loss, we may suffer a reduced ability to predict subsequent anthropogenic impacts on ecosystems worldwide. Given that funding and time are limited, biodiversity research should be focussed on certain taxa for scientifically justified reasons, rather than because of an underlying subjectivity in what we consider to be important. Crucially, conservationists need to be more aware of these unequal weightings to prevent biodiverse taxa being overlooked or understudied.
Redressing biases
Significant challenges remain in addressing the biases we found. One is to popularize these lesser-known taxa to allow recognition of their importance. This could be achieved through more targeted funding for these invertebrate groups (and under-represented countries). Another challenge is to ease the practical issues of identification and research on these taxa [ 33 ]. Opportunities may be found in novel techniques such as metagenomic sequencing [ 34 ], or the development of apps that aid easy identification worldwide [ 35 ]. The use of modern media may ease access to specimens digitally, and help to put researchers and taxonomic experts in touch. It will require a concerted effort to redress these research biases and to ensure the least studied taxa and countries do not remain so, thus ensuring that we maximise the contribution of biodiversity research to our understanding of nature, and minimise the further erosion of biodiversity in our increasingly imperiled world.
Supporting information
S1 dataset. data used in this study..
See ‘metadata’ sheet for more information.
https://doi.org/10.1371/journal.pone.0189577.s001
Acknowledgments
We would like to thank Xavier Bonnet and another anonymous reviewer for their very helpful comments on the manuscript.
- 1. Wilson EO. Biodiversity. National Academies Press; 1988.
- 2. Google [Internet]. 2016 [cited 16 Aug 2016]. Available: https://www.google.co.uk/#q=biodiversity
- 3. Web of Science [Internet]. 2016 [cited 6 Feb 2016]. Available: https://apps.webofknowledge.com
- 4. United Nations. Convention on Biological Diversity [Internet]. 1992. https://doi.org/10.1146/annurev.ento.48.091801.112645
- 5. Gaston KJ. Biodiversity. In: Sodhi N, Ehrlich PR, editors. Conservation Biology for All. Oxford University Press; 2010. https://doi.org/10.1093/acprof:oso/9780199554232.001.0001
- 6. May RM. The dimensions of life on Earth. In: Raven PH, Williams T, editors. Nature and Human Society: the Quest for a Sustainable World. Washington: National Academy; 2000. pp. 30–45.
- 7. Grimaldi D, Engel MS. Evolution of the Insects. Cambridge University Press; 2005.
- View Article
- Google Scholar
- PubMed/NCBI
- 17. RANDOM.ORG [Internet].
- 18. Core Team R. R: A Language and Environment for Statistical Computing [Internet]. Vienna, Austria: R Foundation for Statistical Computing; 2013. Available: http://www.r-project.org/
- 19. IUCN. Table 5: Threatened species in each country (totals by taxonomic group) [Internet]. 2016 [cited 14 Mar 2016]. Available: http://cmsdocs.s3.amazonaws.com/summarystats/2016-2_Summary_Stats_Page_Documents/2016_2_RL_Stats_Table_5_CORRECTED.pdf
- 20. IUCN. Table 1: Numbers of threatened species by major groups of organisms (1996–2014) [Internet]. 2014 [cited 14 Mar 2016]. Available: http://cmsdocs.s3.amazonaws.com/summarystats/2016-2_Summary_Stats_Page_Documents/2016_2_RL_Stats_Table_1.pdf
Academia.edu no longer supports Internet Explorer.
To browse Academia.edu and the wider internet faster and more securely, please take a few seconds to upgrade your browser .
Enter the email address you signed up with and we'll email you a reset link.
- We're Hiring!
- Help Center

Conclusions: Biodiversity Research and Conservation Opportunities

2019, Biodiversity of Angola
Angola is a country full of opportunities. Few countries offer more exciting prospects for young scientists to discover and document the rich biodiversity, complex ecosystem processes and undescribed species of plants and animals that are to be found in its amazing diversity of landscapes and seascapes. The current expanding support of the Angolan government and of international partners is unprecedented, and the positive response from young students ensures the growth of a new generation of biodiversity researchers and conservation professionals. Based on a synthesis of biodiversity research and conservation activities of the past century, we outline opportunities, approaches and priorities for a strengthened collaborative research and conservation agenda.
Related Papers
Biodiversity of Angola
Herbario Lubango
francisco M P gonçalves
Estrela Figueiredo
Plant Ecology and Evolution
Macuntima Pedro
Background and aims – Located in the transition zone of Guineo-Congolian and Zambezian phytochoria, the Angolan provinces of Uíge and Cuanza Norte are of particular interest for floristic studies and high biodiversity is expected. Nevertheless, explorations of the vegetation are relatively rare. Our study aims to supplement a recent checklist of vascular plants of Angola. Methods – Data were collected during 17 field trips between 2013 and 2018, during which herbarium specimens were prepared for later identification of plant species. The results were compared with the current checklist as well as with other floristic works, herbarium collections and online databases relevant for the region.Key results – We document 20 new records of indigenous vascular plant species for the flora of Angola (19 species and one subspecies), including four new generic records. Furthermore, nine alien species are added to the checklist of the flora of Angola.Conclusion – Our results confirm that the flo...
Norbert Jürgens
Angola is a tropical country with many biogeographical units and, therefore, has a high floristic diversity. Although an increasing number of floristic studies has been carried out in Angola in recent years, the country is still considered to be underinvestigated as many species being collected were previously unknown there. Several scientific groups working in different parts of Angola contributed to this paper their data from biodiversity assessments. With this we can add 67 species newly recorded for Angola, including two new generic records and five alien species, to the almost 7,300 vascular plant taxa known so far for Angola. Most of the new records for Angola are also present in different neighbouring countries, but they are little known, and their IUCN threat status has not been assessed yet. However, ongoing fieldwork and exploration are needed to complete the floristic knowledge of the understudied country.
Vladimir Russo
Louis Jacobs
Iain Darbyshire
Biodiversity and Conservation
Mwangi Githiru
Andrzej Gołąbek
This paper presents the use of scaling and dimensional analysis to assess the viability of conventional modelling of immiscible displacement occurring when water is injected into the oil-saturated, porous rock—a conventional secondary oil-recovery method. A brief description of the laboratory tests of oil displacement with water performed on long core sets taken from wells operating on a Polish oil reservoir was presented. A dimensionless product generator based on dimensional analysis and Buckingham Π theorem was used to generate all possible combinatorial sets of dimensionless products for physical variables describing the phenomenon. The mathematical model of the phenomenon was transformed to its dimensionless form, using a selected set of the products. The results of the laboratory tests were analyzed as functions of the products. Statistically verified quantities describing both dependent and independent experiment variables were subject to a regression analysis to study depend...
RELATED TOPICS
- We're Hiring!
- Help Center
- Find new research papers in:
- Health Sciences
- Earth Sciences
- Cognitive Science
- Mathematics
- Computer Science
- Academia ©2024

An official website of the United States government
The .gov means it’s official. Federal government websites often end in .gov or .mil. Before sharing sensitive information, make sure you’re on a federal government site.
The site is secure. The https:// ensures that you are connecting to the official website and that any information you provide is encrypted and transmitted securely.
- Publications
- Account settings
Preview improvements coming to the PMC website in October 2024. Learn More or Try it out now .
- Advanced Search
- Journal List
- Ann Ib Postgrad Med
- v.17(1); 2019 Jun
LOSS OF BIODIVERSITY: THE BURGEONING THREAT TO HUMAN HEALTH
"We are losing biodiversity globally at an alarming rate, and we need a cornucopia of different plants and animals, for the planet's health and our own." 1 Daine Ackerman American poet (1948- )
INTRODUCTION
One of the greatest attributes of the Earth is the biodiversity of her ecosystem. The Convention on Biological Diversity (Article 2) defined biological diversity or biodiversity" as the variability among living organisms from all sources including, inter alia, terrestrial, marine and other aquatic ecosystems and the ecological complexes of which they are part; this includes diversity within species, between species and of ecosystems" . 2 This is a reflection of the more than 9 million types of living thing which include fungi, bacteria, plants, animals, protists that co-inhabit the Earth with human beings and serves as the bases of the diverse array of organismal, genetic and ecological diversity obtainable on Earth. 3 The complex interplay between this rich array of Earth's inhabitants underpins the proper functioning of the Earth's ecosystem. However, the last few decades, have witnessed a rapid disruption and loss of this rich biodiversity in terms of numbers and variability. Human beings are relentless than ever in eliminating genes, species, and biological traits that constitute the biodiversity via her disruptive activities on the Earth ecosystem.
Effects of biodiversity loss
The pressure from agriculture requirement for crop & animal husbandry has made humanity to convert wetlands, forest, and grassland into farmlands and grazing land, among others. Furthermore, the increasing world human population, which has doubled between 1970 and now to more than 7 billion is the other edge of the sword aggravating the global loss of biodiversity. On the other side are factors are exploitation of mineral resources, pollution, the introduction of exotic species & genetically modified organisms, climate changes and alteration and loss habitats which are all connected with human efforts to care for the growing habitats of the Earth.
Every year, at least a species goes into extinction while many species of plants and animals face extinction across the world according to the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services Report (2019). 4
Sadly, most of the global loss of biodiversity occurs in the Developing World, Nigeria inclusive. 5 Furthermore, the critical 25 hotspots of the global loss of biodiversity include areas spanning the rain forest belt of southern Nigeria although the enormous swathe of territory includes the Tropical Andes in South America and Indo-Burma areas in South East Asia. These hotspots are home to a considerable proportion of Earth's species of plants and animals. The Amazon in the Tropical Andes alone harbours 50,000 species or one-sixth of the Earth's total. 6 Generally, known species are going extinct, 1000 times more than newly discovered ones. 7
The history of medicine and the development of the rich armament of pharmacopoeia to combat diseases are traceable to a great deal to derivatives from flora and fauna whether aquatic or terrestrial in origin that are available across the world from ancient time to the present time. This product includes the ubiquitous aspirin derived from the bark of the willow tree Salix alba , which is no more a mere analgesic rather a useful drug in cardiovascular and haematologic disorders treatment. 8 Digoxin, a great drug in managing heart failure is derived from Foxglove Digitalis purpurea L . 8 Drugs such as quinine derived from Cinchona succirubra , and Artemisin based compounds are made from Artemisia annua and have help combat malaria at various times are derived from plant products. 8 While penicillin derived from Penicillium notatum may not be prominent antibiotics in the last 20 years, the subsequent and related products such as Erythromycin are derived from a microorganism. 8 Other great products that have saved many lives include antimitotic agents such as Doxorubicin from Torreya taxifolia and paclitaxel derived from Pacific Yew Taxus brevifolia . The list of products that have been derived from flora and fauna to help combat human disease is endless.
It is interesting to note that only an estimated 10% of the diverse species on Earth have been exploited to combat diseases while a little over 12.5% of the approximately 250 000 species of higher plants have been exploited in the same line. 9 On the other side, only about 1 % of microbial species on Earth are known. 9 , 10
No doubt, the loss of biodiversity does not only threaten new drug discovery especially in the light of emerging and reemerging diseases, but it also threatens the ability to discover a more effective therapy for the burgeoning non-communicable diseases, hence man's quest to stem the tide of this increasing global burden.
Equally threatened is the accessibility to clean fresh water and good quality air. 8 The air and water are greatly more polluted than ever due to human industrial activities, while the purifying capacity of the ecosystem is being lost. 2
Food production is being affected, thereby endangering the nutritional status of the world population, especially in regions where the poorest habit. The economic activities of natural pollinators, e.g. bees which aids our plants to fruits are estimated at approximately $550billion. 11 The aquatic species are being depleted. Fisheries currently provide 16% of the global protein source. The wild flora and fauna could mean alternative access to the nutritious source to the aforementioned is being wiped out.
While the biodiversity is contracting, there is a great danger of emerging and reemerging of infectious agents which threatens global health. The incidence of Lyme disease or West Nile virus (WNV) in humans have been linked to the loss of biodiversity among the animal host. 12
While the loss of biological biodiversity appears to affect significantly human health, it has also been opined to be a significant threat to the attainment of sustainable development goals which is the blueprint for achieving a better and more sustainable future for all. 13 Currently it threatens the goal 1; to reduce poverty, Goal 2; zero hunger, Goal 3; good health & well-being and goal 6; clean water and sanitation, Goal 11; sustainable cities and communities, Goal 13; Climate Action, Goal 14; Life Below Water and Goal 15; life on land. 14
RECOMMENDATIONS
While it may be easier to pass the attempt at the reversal of this loss to the tuft of core conservationists it is necessary to point out that biomedical scientists or researchers and even clinicians who will prescribe the yet undiscovered drugs from flora or fauna have an enormous role to play. In the same vein, the temptation to isolate this discussion to only the conservationist circle, may not be optimal to achieve broad-spectrum coalition for advocacy. Now is the time biomedical scientists join the advocacy to stem the tide of biodiversity loss by joining the luxurious pool necessary for such advocacy.
Conservative efforts should be in the mainstream of discourse in scientific fora of biomedical scientists since this loss threatens the whole capability of modern medicine to keep confronting the burgeoning disease burden.
Furthermore, the least maybe in our local environment we do is to promote and join conservation efforts such as serving as an advocate for the protection of endangered species, increase in protected areas of flora and fauna and exercise high ethics in animal researches involving endangered species.
Indeed, a little there, little here approach may help save the world. A tree or other exotic nurtured in an enforced green space in a rural clinic may be in a right direction and commendable effort at mitigating this ensuing disaster.
There is a need for more interdisciplinary research on the interrelationship of biodiversity loss with medical and medical-related themes. There is also a greater need for inquiries into the usefulness of the numerous natural products in solving human disease miseries. It may be the molecule to cure some of the vast arrays of presently incurable diseases are in some species in the Niger Delta area or Amazon.
Finally, there is a need for the national governments to enact and implement laws to protect the endangered species like the United States Endangered Species Act (1973). It would also be necessary to advocate for domestication locally the Aichi Biodiversity Targets proposed at the Convention on Biological Diversity (CBD) in Nagoya, Japan, in October 2010, which target 1 explicitly state "By 2020, at the latest, people are aware of the values of biodiversity and the steps they can take to conserve and use it sustainably" . 4 , 15
Although we may not be able to save the extinct species with their lockup potentials, we may at least be able to wrestle the endangered & threatened species from similar extinction. Humanity needs to live a life accommodative of other species to promote healthy and robust dynamism among species, thereby sustaining healthy lives for ourselves and healthy interactions with the ecosystems. The ultimate goal, therefore, will be to halt the rate of biodiversity loss and ensure a stable ecosystem as soon as possible.
Biodiversity Research Paper

This sample diversity research paper on biodiversity features: 4800 words (approx. 16 pages) and a bibliography with 12 sources. Browse other research paper examples for more inspiration. If you need a thorough research paper written according to all the academic standards, you can always turn to our experienced writers for help. This is how your paper can get an A! Feel free to contact our writing service for professional assistance. We offer high-quality assignments for reasonable rates.
It is certain that the human species has to accept sharing the Earth and its ecosystem not only with other humans but also with other living beings.
Academic Writing, Editing, Proofreading, And Problem Solving Services
Get 10% off with 24start discount code.
Human activities can cause irreversible changes that harm this ecosystem. Human activities, justifiable as they may be for survival or development of one group, when harmful can lead to excesses, neglecting duties to other people or other species as well as the environment. Interdependence of humans and other living beings must be addressed for the survival of all species. The human species, having a predominant position within the biosphere, has a duty to care for the Earth and its biosphere, not as its owner but rather as its manager; this means not considering only producing immediate gains but also sustaining the vital interests of our species, in present and future generations. Solutions that balance immediate and long-term benefits may be found using a global perspective, taking account of all interlinked parameters in the world system: economic growth, consumption patterns, lifestyles, scientific and technological progress, and above all justice and equity. This necessitates addressing problems in the framework of international cooperation, in the spirit of universal solidarity. There are therefore multiple ethical aspects to biodiversity.
Introduction
Since the last century, in just a few decades, concern about an increasing rate of loss of biodiversity has become widespread among the public. It has ceased to be an exclusive concern of well-informed persons, ecology or biology scientists and experts. Indeed, these losses have been examined in the media, and it is becoming common knowledge that the loss of biodiversity is intimately linked to ecosystem changes, some of which are already visible and tangible. In places, ordinary people are already directly or indirectly coping and adapting to the global atmosphere’s warming, which is a major change in the global ecosystem. In places, people are facing historically unpredictable flooding of formerly inhabited lands or islands; in other places people are facing severe droughts, the spreading of deserts and consequent famine, shortage of clean drinking water, or unexpected health problems. These local upheavals can be causally linked to the change in the global ecosystem, and increasing rate of loss of biodiversity is another result. People are experiencing these changes over the span of one lifetime; significant changes and loss of biodiversity are in some places observed and experienced not in terms of millenniums or centuries but rather over decades or years.
This research paper will outline the concept of biodiversity, its meaning, the range of its dimensions, and its implications notably from an ethical point of view.
History And Development Of The Concept
Over the ages, Homo sapiens of all cultures have observed the variability of the living organisms in nature – countless varieties of plants with leaves and flowers of different colors and scents, all sorts of fruits with various flavors, and so much variety among animals: some crawling, some walking, some flying, and some swimming.
The fact that vital resources (e.g., food, medicine, and other services) are derived from nature has inevitably convinced humans, even by intuitive understanding, that variety in living nature has importance and interest for human existence. Indeed, humans must always have intuited that for survival, diversity in living nature is advantageous. That is the source for the idea that, given human capacity to do harm, there is now a duty that the diverse ecosystem should be protected. Yet, it is possible that humans could mistakenly believe that the healthy ecosystem, which has always been there, will remain stable and unchanged for all time.
The fragility of the ecosystem was foreseen by some philosophers of antiquity like Lucretius (98–54 BC) in his poetic work “De rerum natura” and its diversity recorded by scientists of the nineteenth century. Indeed, the present assertion that the diversity of living nature is to be protected from overexploitation is due to relatively recent progress of scientific knowledge. In particular it is the theory of evolution, the basis of modern biology, that explains that a healthy ecosystem is not stable in an unchanging way. By careful observation and deduction, it is now understood that over history, some living species have declined in number, and some become extinct. After Jean-Baptiste Lamarck completed the classification of invertebrates, Charles Darwin published in the year 1859 the first edition of On the Origin of Species (Darwin 1872), in which he highlights as a principle that these classifications of living nature are not fixed and stable. He was the first to formulate a reliably verifiable argument for evolution of species, by means of the operation of natural and sexual selection. Darwin upholds the thesis that living species are not fixed in groups and that they diversify over generations. Over time a particular species or population may increase in number or decline and even disappear. To explain changes that gradually arise and spread in a population, Darwin’s concept of natural selection allowed us to recognize how success of some traits is conditioned by interaction with the natural environment. Thanks to the work on genetics by Mendel, this theory of evolution is even better understood today. Evolution describes, within a given population, change in the inherited characteristics over successive generations toward those favored by a given ecosystem. An ecosystem can condition and give rise to diversity at every level of biological organization, including species, individual organisms, and molecules such as DNA and proteins (Bowler 1989). As a system, it will also be more stable if it can withstand some loss of biodiversity (as may occur in a drought year), if populations that remain are themselves diverse enough to “replace” the losses, as they diversify with time. Indeed, the most resilient ecosystem will necessarily include a great diversity of life forms or “biodiversity.”
Scientists agree that present biodiversity losses are due to multiple factors, some natural and some attributable to human activities (ICSU 1992). According to the political concept of conservation of nature, societies have to prepare to conserve the nature’s diversity by various measures, including by ensuring that any human exploitation of nature is justifiable, equitable, and ecologically sustainable. For instance, logging today causes vast areas to be deforested, leading also to desertification; industrial-scale agriculture today results in disrupted virgin forests and the invasion by nonnative species that establish and spread outside their normal habitat; harsh exploitation of land to extract raw materials on the surface, underground, at sea bottom, or from lakes or other waters may be justified by our present human development needs. But is the destructiveness of the practices used really necessary? Multiplying destructive practices can certainly harm ecosystems and cause the loss of biodiversity. Biodiversity plays an important role in the way ecosystems function, and drastic loss of biodiversity will impede the multitude of services people obtain from these ecosystems: nutrients, water recycling, soil formation and retention, resistance against invasive species, pollination of plants, regulation of climate, as well as pest and pollution control.
Ecosystem changes have positive or negative influence on biodiversity loss/gain rates in the immediate sense, for example, when atmosphere become warmer in the context of climate change (Meakin 1992). Scientists have contributed to an understanding that some human activities contribute, and possibly exacerbate, to ongoing environmental changes, and the rate and speed of biodiversity loss has come to be used as indication of a negative influence. It is now understood that human activities have increased the extinction species rate by at least 100 times compared to a natural (prior) rate. Confronted with the decline and extinction of some species, political conscience has been raised all over the world for improving the protection of nature. In some places, nature is being viewed as part of the shared commons or shared patrimony. Especially in the case of wasting natural resources, there is great concern. The conservationist idea has gradually led to measures to protect spaces like national parks, to protect the quality of air and water, and to establish use controls, such as for hunting and fishing or for land use. In parallel, relatively recent progress in the field of genetic engineering has allowed spread and commercialization of genetically modified organisms (GMOs). These novel life forms that did not previously occur in nature are thus being introduced, in agriculture, for example. This gives rises to important questions not only for the health of the first consumers but also and especially for the long-term impacts on ecosystems on which we depend. Increased political sensitivity for conservation of ecosystems may help to measure for long-term impact monitoring and control even in this area.
The twentieth century saw many persons resolutely rally for conservation, as a worldwide social movement was born. From the first international conference for the protection of nature held in Berne (1913) to the world conservation congress (Korea, 2012) held by the International Union for Conservation of Nature, this was a century of action, aiming to achieve reduction in the rate of biodiversity losses. Indeed, many reports have emphasized that human actions often load to irreversible losses in terms of diversity of life on Earth, and these losses have been more rapid in recent decades than ever before in human history. By the year 1968, the United Nations Educational, Scientific, and Cultural Organization (UNESCO) established the Man and Biosphere (MAB) group with the role of promoting an intergovernmental research program to provide the scientific basis for a sustainable management of natural resources in an approach that balances ecological, social, and economic concerns. Some years later, the project “The limits to growth” was launched, aiming to alert political and media decision-makers to the seriousness of environmental problems, notably to highlight the need to balance rapid economic growth against ecological limits. At the same time, the United Nations Environmental Program (UNEP) was established as a result of the first Earth Summit held in Stockholm (1972). By 1980, the concerns relating to the impacts of human activities on species and ecosystems give rise to a new branch of biology, called the “biology of conservation,” meant to support the implementation of measures and actions for conserving nature.
The Earth Summit of Rio, held in 1992 at Rio de Janeiro (Brazil), could be considered as the turning point when for the first time a shared realization that environmental change is posing a threat to the habitability of the planet Earth could support a consensus for taking political action. The Earth Summit convened representatives from 178 nations, nongovernmental agencies, and many other interested parties among which thousands of members of the media; and it focused on global environmental issues that would become central to policy implementation. During this meeting, delegates adopted the Convention on Biology Diversity (CBD) marking a collective effort of wills, recognizing that sustainable development and the protection of biodiversity must be considered as a common concern of humanity. The CBD has become the framework of national strategies to preserve biodiversity (Meakin 1992).
After the Earth Summit of Rio, the parties to the CBD agreed to achieve the reduction of the current biodiversity loss by 2010. However, the target assessment has shown that the biodiversity losses and the related changes in the environment are now faster than ever before, and there is no sign of this process slowing down. Notwithstanding the best intentions of human race to live responsibly as demonstrated at Rio, uneven and insufficient progress in sustainable development was shown 20 years later, during the United Nations Conference on Sustainable Development (UNCSD 2012, otherwise named Rio + 20). It is likely that progress on sustainable development is slow because the global political system is not working smoothly on a matter of this scale and complexity. Indeed, multiple interlinked factors have to be taken into account sustainable development practices to develop, and practices adopted in all parts of the world: notably, assessments and decisions must integrate economic, social, and environmental aspects and support transition to sustainable practices. Indeed, at the Rio + 20 summit, heads of state recognized this complexity, specifically noting “a need to achieve sustainable development by promoting sustained, inclusive and equitable economic growth, creating greater opportunities for all, reducing inequalities, raising basic standards of living, fostering equitable social development and inclusion, and promoting integrated and sustainable management of natural resources and ecosystems that supports, inter alia, economic, social and human development while facilitating ecosystem conservation, regeneration and restoration and resilience in the face of new and emerging challenges” (UNCSD 2012) – challenges such as financial crises, food and/or water security crises, or forced migration.
Conceptual Clarification/Definition
Etymologically, the term biodiversity is made up of two roots, that is to say biology (from Greek bios = life) and diversity, contracted to biodiversity. The term is in itself seemingly obvious and is largely used even by ordinary mortals, each one with his understanding. And yet, this term is proving to be a complex concept.
At first sight, the concept biodiversity refers to the number of species of animals, plants, and microorganisms in the nature, in other words specific biodiversity, species being the most evident element as it corresponds to the basic unit of a classification of living beings. Taxonomy has counted about 1.7 million species, probably just the most visible part of biodiversity. Recent studies estimate the total number of species between three and ten million. Many species are not very accessible for study, including unicellular organisms and bacteria. The marine biodiversity is still in large part unknown because about 95 % of oceans remain unexplored.
Furthermore, within species there is an enormous variability of genes between individuals as well as populations. This means there is genetic diversity within a same species which is essential to confront to environmental change by allowing the population’s adaptation through evolution.
Living beings adapt to different environments everywhere: on land such as deserts, rainforests, or underground galleries and in water as ocean, lakes, ponds, or other watercourses. It is in these environments, also called ecosystems, that living populations interact with each other and all that is around. So, ecosystems draw on biological diversity.
Biodiversity is exhaustively defined, in the article 2 of the Convention on Biological Diversity: Biodiversity or biological diversity means the variability among living organisms from all sources including, inter alia, terrestrial, marine and other aquatic ecosystems and ecological complexes of which they are part: this includes diversity within species, between species and ecosystems (CBD 1992). The concept refers also to the presence of humans who depend on biodiversity for sustainable development of societies. So, biodiversity exists at three levels of organization: genes, species, and ecosystems; biodiversity is not limited to describing the variety of species but represents also interactions between living beings and also between them and their physical and chemical environments at different levels. The term covers how this diversity changes from one location to another over time. Indicators such as the number of species in a given area can help in monitoring some aspects of biodiversity but remain incomplete and insufficient for providing an accurate picture of the extent and distribution of all components of biodiversity. History has it that the expression biological diversity was coined by Thomas Lovejoy. This conservation biologist introduced the term to the scientific community in 1980. It was then used by Walter Rosen during a congress entitled “The National forum on Biodiversity” held in Washington in 1986.
Ethical Dimensions
Human beings within nature, the unity of the earth’s ecosystem.
It is said that in 1854, a man named Seattle, representing several Indian (first inhabitants) tribes, replied to pressure from the US government to buy Indians’ land, by an ecological appeal that still resounds today in a universal echo. He reportedly said “We are part of this land; this land is part of us… Rivers are our sisters, they quench our thirst, carry our canoes and feed our kids… the sweet-scented flower is our sister; stag, horse, the big eagle are our brothers; the rocky crest, the sap in meadow, the heat of pony, and human; all are members of the same family. .. .When all buffalos will be slaughtered, wild horses tamed, the secret nooks of forest overload with the odor of many humans, and the view of flowered hills tarnished by the speaking wires… Then, where will be thickets? Disappeared. Where will be eagle? Extinct. And that extinction will mark the end of life, and the beginning of survival.”
The consensus of scientists is that the theory of evolution is one of the most reliably established facts. Evolution means not only change, but also it implies continuity. Each species, no matter how small, has an important role to play in a web of life. Species depend on each other. Even while there might be “survival of the fittest” within a given species, each species depends on the services provided by other species to ensure its survival. It is a type of cooperation based on mutual survival (a balanced ecosystem often refers to this cooperation). Studies on the field of virgin ecosystems tend to show that a healthy or balanced ecosystem can better withstand and recover from a variety of disasters. Indeed, ecosystems – if left alone – appear to have capacity to self-sustain. Viewed from this angle, biodiversity has an intrinsic value; it should be respected, regardless of being useful for humans. Ultimately, concern for the level of biodiversity on Earth is to be justified not just in relation with its utility but also for ethical reasons, despite that some economic methods assign monetary values to benefits (e.g., services such as recreation or clean drinking water).
Human Responsibility
Humans are one component of an ecosystem and often in a predominant position. Indeed, at the present state of our knowledge, humans are the only living beings capable of conscience, self-criticism, and moral judgment. Humans are the only living beings to whom moral responsibility could be attributed. Moreover, in organized human society the capacity of humans is still greater than at the level of one individual.
Human responsibility includes obligations toward future generations. Indeed, the concept of humanity refers to the principle of human dignity as emphasized as well in the Universal Declaration on Human Rights (1948) as in the Universal Declaration on Bioethics and Human Rights (2005). An intrinsic value is recognized in human-kind considered as a whole, not only consisting in each of all individual humans that now exist all over the world but also including future generations. Thus, a moral responsibility is assigned to the present generation: the obligation to bequeath to future generations a planet that remains a place in which it is pleasant to live. To meet that responsibility, humans must not devastate the Earth today.
Recognition of the preeminent place of humans as a species also leads to attributing other moral obligations. Humans have moral obligations toward not only other humans of the future but also toward other living beings living today as well as the biosphere in its entirety. And, in this light, we may have now to review our management and action methods especially on pertaining to the other species with whom we coexist. The best understanding of the role of humans could be as a manager, a kind of primus inter pares, with a preeminent place and responsibility related to biodiversity. Humans then have responsibility to manage carefully the transformation of nature. Indeed, humans affect the balance of natural processes. Disrupting nature is also part of human nature. The Universal Declaration on Bioethics and Human Rights recognizes that humans have a special responsibility to protect biodiversity and the biosphere within which human beings exist (UNESCO 2009).
To yearn for well-being, welfare, good health, and comforts are, among others, legitimate motivations to justify most of human activities. It is, however, demonstrated that the interest of humanity and human health is also dependent on a pleasant environment; that means to safeguard the welfare of the biological species and their ecosystems. The ethical principle of beneficence and non-malfeasance, as stipulated in the article 4 of the Universal Declaration on Bioethics and Human Rights, implies the imperative to tolerate and manage acknowledging this beneficial interdependence (UNESCO 2009).
Biodiversity, A Complex And Global Challenge
A holistic paradigm.
The ethical questions posed at the present stage of history, all over the world, are sufficiently complex that they do not suggest a holistic paradigm. It is a challenge to identify at first a cohesive view. Maintaining biodiversity is unquestionably a problem of multidisciplinary nature and wide concern. Problems may in some cases be solved by unexpected compromises between interests that seemingly diverge. Maintaining biodiversity is a global challenge that entails questions such as justice and equity, solidarity and cooperation, virtue of moderation in the wild race for profit, economic growth, sustainable development, the common interest of humanity (the present as well as the future generation), etc. The Universal Declaration on Bioethics and Human Rights, although it put great emphasis on humans and responsibilities toward humans, also extends the concern to the rest of living beings. The Earth’s biosphere must function, as a whole, with its constant pursuit of harmony.
Solidarity And Cooperation
The world regions cannot longer be separated and cannot pursue totally independent interests, especially in relation to the ecosystem’s problems. In some cases, ecologic damages caused in a given part of the globe could be harmful at a diametrically opposite spot. Ecosystem changes such as thinning of the ozone layer due to excess of industrialization, global warming, and necessity to conserve vegetation, for the synthesis of oxygen, may have severe consequences in undeveloped regions where people have hardly ever benefit from civilization progresses; rising sea levels are leading to disappearance of some islands that were inhabited before. In fact, atmospheric air, oceans, and other watercourses are like connected vessels for the one same Earth. And really, every inhabitant of the planet is “in the same boat”; all must realize that humans have a common planetary destiny and that there is an inevitable necessity of solidarity for survival, when it comes to giving priority to the biosphere. Sustainable development must be inclusive, benefiting and involving all people.
All countries, developed or developing, landlocked, or small islands, face specific challenges to achieve sustainable development. In the context of national and international political institutions, the challenge is to consider national sovereignty as well as to deal with international imperatives, to treat separately the problems posed in each region, as well as to have in mind integration into global interests. In a pragmatic and flexible way, actions have to be expended having to make the best of the sacrosanct principle of national freedom and sovereignty on the one hand, the imperatives dictated by the vital interests of humanity on the other hand; to find trade-offs reconciling people’s particular historical background to some established realities that can be scientifically anticipated. Eventually, these institutions are likely to impose worldwide strategies. Because it is a question of survival for everybody, there is a basis for a true cooperation, in which everyone may fully participate and contribute, even and especially those who have limited powers. To paraphrase Martin L. King, we have to learn living together as brothers (and sisters); otherwise, we will die together like idiots.
Public Awareness
Debates on biodiversity questions are, with good reason, carried out among intellectuals, scientific and technology experts, taking account of their complexity. However, because solutions depend not only on political decisions but also on the attitudes of those who will have to carry them out in their daily lives, it appears obvious that public awareness of different possible schemes should be increased. The public has to be informed about the benefits of conservation, especially so that it may consider trade-offs between different options; this could help to maximize the benefits to society.
Economic And Social Issues
From an economic point of view, various goals pursued in society to improve business activities depend on biodiversity and modify ecosystems. It has been demonstrated, for example, that actions to increase food production can lead to reduce water availability for other uses with, as a consequence, deterioration in other ecosystem services. The value of services lost may greatly exceed the short-term economic benefits that are gained (UNESCO 1999). It has to be pointed out that a large number of the world’s poor people rely directly on biodiversity and ecosystem services, and then livelihoods would be affected first and foremost by decrease in the services related to food production, nutrition, water, and sanitation. Indeed, over the last century, some people have benefited from the conversion of natural ecosystems and increase in international trade, while others have suffered from the consequences of biodiversity losses and from restricted access to resources they depend upon. Changes in ecosystems are harming many of the world’s poorest people, who are the least able to resist these changes (UN 2014).
Virtue Of Moderation And Ethics Of Care
The consequences of biodiversity loss and reduction in ecosystem services call out to the conscience of each the virtue of moderation. Indeed, any enterprise inevitably leads to nuisances for other sectors if it goes too far. Businesses that aim only at the highest profits and unlimited growth cumulatively result in excesses in the consumption patterns of goods or services and endless innovative technologies consuming ever-increasing raw materials, though these are limited in quantity and increasingly precious. Considering, for example, the controversies raised by GMO production, the GMO developers view this technology as aiming to increase agriculture and livestock yield, enhance nutrition, and provide other benefits for humans; on the other hand, GMOs may be connected with health problems and environmental damage, and, most importantly, the long-term impacts of GMOs are unknown. Indeed, no one, even the biggest chemical companies producing GMOs, can anticipate their future consequences; moreover, they cannot tell if the novel life forms they create, once widely released into the environment, could be recalled if they are proving to be harmful. When actions are taken without considering potential consequences, these will lead inevitably to imbalance. Implementation of an ethics of care is proving to be a vital necessity for all, especially if things are considered in the long term. Direct and indirect drivers of biodiversity loss must be addressed to better protect biodiversity and ecosystem services
Considering things at the planetary scale, it is now assessed that biodiversity losses and its corollary, the deterioration of ecosystem services, are faster in recent decades than ever before in the history of humanity. Even if parts were attributed to natural causes, the damage resulting from human activities can be addressed.
Normal human activities, even legitimate and perfectly justifiable by several human needs like agriculture, livestock, business, and exploiting raw materials for development, become harmful and, at the end, inevitably lead to serious imbalance if they go too far, without taking care not to harm other people or other living species as well as the environment. Ethics of care is proving to be appropriate, with consideration of the long term. Moreover, biodiversity should be attributed an intrinsic value, beyond its utility for humans. So, firstly considered as res nullius, biodiversity gradually appears as having amenity and ethics value.
The Earth is one and common not only for the whole humanity but also for nonhuman living beings, that is to say animals, plants, and microorganisms. Humans have to confront this reality that cannot be ignored. Taking account of the particularities of human capacities from a moral point of view, in comparison with and in relation to other living species, humans have a preeminent position within the biosphere. Humans therefore have a responsibility to carefully manage this Earth, not as the owner but just as the manager. There is an interdependence that must be addressed for survival of all species including humans.
Reflections on issues related to biodiversity and ecosystem balance inevitably lead to consider things at the global level, aiming to harmoniously integrate the interests of many. This work has to be done in universal solidarity, international cooperation, and open, sincere, and honest dialogue, with benevolence and attention to other people, listening even to those who are limited in their power, as well those who have great expertise in scientific, technological, financial, or other domains. The long-term continuation of life on the planet is at stake.
Bibliography :
- Bowler, P. (1989). The Mendelian revolution the emergence of hereditarian concepts in modern sciences and society. Baltimore: John Hopkins University Press. ISBN 0-485-11375-9.
- Convention on Biological Diversity. (1992). About the Convention. https://www.cbd.int/convention/
- Darwin, C. (1872). On the origin of species by mean of natural selection, or the preservation of favoured races in the struggle for life. London: John Murray, 5(121), 318
- (1992). Proceedings of the International Conference on Agenda of science for Environment and Development into 21st Century (ASCENT 21). Paris: ICSU.
- Meakin, S. (1992). Rio Earth Summit 1992. Summary of the United Nations Conference on Environment and Development. https://sustainabledevelopment.un.org/milestones/unced
- (2014). Report of the Open Working Group proposal for Sustainable Development Goals. https://sustainabledevelopment.un.org/
- (2012). Report of the United Nations Conference on Sustainable Development. Rio de Janeiro, 2012. Final document page 8 “The future we want.” https://sustainabledevelopment.un.org/content/documents/733FutureWeWant.pdf
- (1999). Declaration on science and the use of scientific knowledge. Adopted at the World Conference on Science organized by ISCU/Unesco, Budapest, 26 June–1 July 1999. http://www.unesco.org/science/wcs/eng/declaration_e.htm
- (2009). Tandon, P.N. : Protection of the environment, the biosphere and biodiversity. In H. A. M. J. ten Have & M. S. Jean (Eds.), The UNESCO Universal Declaration on Bioethics and Human Rights. Background, principles and application. Paris: UNESCO.
- (2012). Report on the World Conservation Congress. Korea. https://www.iucn.org/
- Maris, V. (2010). Philosophie de la Biodiversité. Petite éthique pour une nature en péril. Paris: Buchet/Chastel.
- (1987). Our common future: Towards sustainable development. Report of the World Commission on Environment and Development “Our Common Future” (Brundtland report). https://sustainabledevelopment.un.org/content/documents/5987our-common-future.pdf
ORDER HIGH QUALITY CUSTOM PAPER

This paper is in the following e-collection/theme issue:
Published on 12.4.2024 in Vol 26 (2024)
Application of AI in in Multilevel Pain Assessment Using Facial Images: Systematic Review and Meta-Analysis
Authors of this article:
- Jian Huo 1 * , MSc ;
- Yan Yu 2 * , MMS ;
- Wei Lin 3 , MMS ;
- Anmin Hu 2, 3, 4 , MMS ;
- Chaoran Wu 2 , MD, PhD
1 Boston Intelligent Medical Research Center, Shenzhen United Scheme Technology Company Limited, Boston, MA, United States
2 Department of Anesthesia, Shenzhen People's Hospital, The First Affiliated Hospital of Southern University of Science and Technology, Shenzhen Key Medical Discipline, Shenzhen, China
3 Shenzhen United Scheme Technology Company Limited, Shenzhen, China
4 The Second Clinical Medical College, Jinan University, Shenzhen, China
*these authors contributed equally
Corresponding Author:
Chaoran Wu, MD, PhD
Department of Anesthesia
Shenzhen People's Hospital, The First Affiliated Hospital of Southern University of Science and Technology
Shenzhen Key Medical Discipline
No 1017, Dongmen North Road
Shenzhen, 518020
Phone: 86 18100282848
Email: [email protected]
Background: The continuous monitoring and recording of patients’ pain status is a major problem in current research on postoperative pain management. In the large number of original or review articles focusing on different approaches for pain assessment, many researchers have investigated how computer vision (CV) can help by capturing facial expressions. However, there is a lack of proper comparison of results between studies to identify current research gaps.
Objective: The purpose of this systematic review and meta-analysis was to investigate the diagnostic performance of artificial intelligence models for multilevel pain assessment from facial images.
Methods: The PubMed, Embase, IEEE, Web of Science, and Cochrane Library databases were searched for related publications before September 30, 2023. Studies that used facial images alone to estimate multiple pain values were included in the systematic review. A study quality assessment was conducted using the Quality Assessment of Diagnostic Accuracy Studies, 2nd edition tool. The performance of these studies was assessed by metrics including sensitivity, specificity, log diagnostic odds ratio (LDOR), and area under the curve (AUC). The intermodal variability was assessed and presented by forest plots.
Results: A total of 45 reports were included in the systematic review. The reported test accuracies ranged from 0.27-0.99, and the other metrics, including the mean standard error (MSE), mean absolute error (MAE), intraclass correlation coefficient (ICC), and Pearson correlation coefficient (PCC), ranged from 0.31-4.61, 0.24-2.8, 0.19-0.83, and 0.48-0.92, respectively. In total, 6 studies were included in the meta-analysis. Their combined sensitivity was 98% (95% CI 96%-99%), specificity was 98% (95% CI 97%-99%), LDOR was 7.99 (95% CI 6.73-9.31), and AUC was 0.99 (95% CI 0.99-1). The subgroup analysis showed that the diagnostic performance was acceptable, although imbalanced data were still emphasized as a major problem. All studies had at least one domain with a high risk of bias, and for 20% (9/45) of studies, there were no applicability concerns.
Conclusions: This review summarizes recent evidence in automatic multilevel pain estimation from facial expressions and compared the test accuracy of results in a meta-analysis. Promising performance for pain estimation from facial images was established by current CV algorithms. Weaknesses in current studies were also identified, suggesting that larger databases and metrics evaluating multiclass classification performance could improve future studies.
Trial Registration: PROSPERO CRD42023418181; https://www.crd.york.ac.uk/prospero/display_record.php?RecordID=418181
Introduction
The definition of pain was revised to “an unpleasant sensory and emotional experience associated with, or resembling that associated with, actual or potential tissue damage” in 2020 [ 1 ]. Acute postoperative pain management is important, as pain intensity and duration are critical influencing factors for the transition of acute pain to chronic postsurgical pain [ 2 ]. To avoid the development of chronic pain, guidelines were promoted and discussed to ensure safe and adequate pain relief for patients, and clinicians were recommended to use a validated pain assessment tool to track patients’ responses [ 3 ]. However, these tools, to some extent, depend on communication between physicians and patients, and continuous data cannot be provided [ 4 ]. The continuous assessment and recording of patient pain intensity will not only reduce caregiver burden but also provide data for chronic pain research. Therefore, automatic and accurate pain measurements are necessary.
Researchers have proposed different approaches to measuring pain intensity. Physiological signals, for example, electroencephalography and electromyography, have been used to estimate pain [ 5 - 7 ]. However, it was reported that current pain assessment from physiological signals has difficulties isolating stress and pain with machine learning techniques, as they share conceptual and physiological similarities [ 8 ]. Recent studies have also investigated pain assessment tools for certain patient subgroups. For example, people with deafness or an intellectual disability may not be able to communicate well with nurses, and an objective pain evaluation would be a better option [ 9 , 10 ]. Measuring pain intensity from patient behaviors, such as facial expressions, is also promising for most patients [ 4 ]. As the most comfortable and convenient method, computer vision techniques require no attachments to patients and can monitor multiple participants using 1 device [ 4 ]. However, pain intensity, which is important for pain research, is often not reported.
With the growing trend of assessing pain intensity using artificial intelligence (AI), it is necessary to summarize current publications to determine the strengths and gaps of current studies. Existing research has reviewed machine learning applications for acute postoperative pain prediction, continuous pain detection, and pain intensity estimation [ 10 - 14 ]. Input modalities, including facial recordings and physiological signals such as electroencephalography and electromyography, were also reviewed [ 5 , 8 ]. There have also been studies focusing on deep learning approaches [ 11 ]. AI was applied in children and infant pain evaluation as well [ 15 , 16 ]. However, no study has focused on pain intensity measurement, and no comparison of test accuracy results has been made.
Current AI applications in pain research can be categorized into 3 types: pain assessment, pain prediction and decision support, and pain self-management [ 14 ]. We consider accurate and automatic pain assessment to be the most important area and the foundation of future pain research. In this study, we performed a systematic review and meta-analysis to assess the diagnostic performance of current publications for multilevel pain evaluation.
This study was registered with PROSPERO (International Prospective Register of Systematic Reviews; CRD42023418181) and carried out strictly following the PRISMA (Preferred Reporting Items for Systematic Reviews and Meta-Analyses) guidelines [ 17 ] .
Study Eligibility
Studies that reported AI techniques for multiclass pain intensity classification were eligible. Records including nonhuman or infant participants or 2-class pain detection were excluded. Only studies using facial images of the test participants were accepted. Clinically used pain assessment tools, such as the visual analog scale (VAS) and numerical rating scale (NRS), and other pain intensity indicators, were rejected in the meta-analysis. Textbox 1 presents the eligibility criteria.
Study characteristics and inclusion criteria
- Participants: children and adults aged 12 months or older
- Setting: no restrictions
- Index test: artificial intelligence models that measure pain intensity from facial images
- Reference standard: no restrictions for systematic review; Prkachin and Solomon pain intensity score for meta-analysis
- Study design: no need to specify
Study characteristics and exclusion criteria
- Participants: infants aged 12 months or younger and animal subjects
- Setting: no need to specify
- Index test: studies that use other information such as physiological signals
- Reference standard: other pain evaluation tools, e.g., NRS, VAS, were excluded from meta-analysis
- Study design: reviews
Report characteristics and inclusion criteria
- Year: published between January 1, 2012, and September 30, 2023
- Language: English only
- Publication status: published
- Test accuracy metrics: no restrictions for systematic reviews; studies that reported contingency tables were included for meta-analysis
Report characteristics and exclusion criteria
- Year: no need to specify
- Language: no need to specify
- Publication status: preprints not accepted
- Test accuracy metrics: studies that reported insufficient metrics were excluded from meta-analysis
Search Strategy
In this systematic review, databases including PubMed, Embase, IEEE, Web of Science, and the Cochrane Library were searched until December 2022, and no restrictions were applied. Keywords were “artificial intelligence” AND “pain recognition.” Multimedia Appendix 1 shows the detailed search strategy.
Data Extraction
A total of 2 viewers screened titles and abstracts and selected eligible records independently to assess eligibility, and disagreements were solved by discussion with a third collaborator. A consentient data extraction sheet was prespecified and used to summarize study characteristics independently. Table S5 in Multimedia Appendix 1 shows the detailed items and explanations for data extraction. Diagnostic accuracy data were extracted into contingency tables, including true positives, false positives, false negatives, and true negatives. The data were used to calculate the pooled diagnostic performance of the different models. Some studies included multiple models, and these models were considered independent of each other.
Study Quality Assessment
All included studies were independently assessed by 2 viewers using the Quality Assessment of Diagnostic Accuracy Studies 2 (QUADAS-2) tool [ 18 ]. QUADAS-2 assesses bias risk across 4 domains, which are patient selection, index test, reference standard, and flow and timing. The first 3 domains are also assessed for applicability concerns. In the systematic review, a specific extension of QUADAS-2, namely, QUADAS-AI, was used to specify the signaling questions [ 19 ].
Meta-Analysis
Meta-analyses were conducted between different AI models. Models with different algorithms or training data were considered different. To evaluate the performance differences between models, the contingency tables during model validation were extracted. Studies that did not report enough diagnostic accuracy data were excluded from meta-analysis.
Hierarchical summary receiver operating characteristic (SROC) curves were fitted to evaluate the diagnostic performance of AI models. These curves were plotted with 95% CIs and prediction regions around averaged sensitivity, specificity, and area under the curve estimates. Heterogeneity was assessed visually by forest plots. A funnel plot was constructed to evaluate the risk of bias.
Subgroup meta-analyses were conducted to evaluate the performance differences at both the model level and task level, and subgroups were created based on different tasks and the proportion of positive and negative samples.
All statistical analyses and plots were produced using RStudio (version 4.2.2; R Core Team) and the R package meta4diag (version 2.1.1; Guo J and Riebler A) [ 20 ].
Study Selection and Included Study Characteristics
A flow diagram representing the study selection process is shown in ( Figure 1 ). After removing 1039 duplicates, the titles and abstracts of a total of 5653 papers were screened, and the percentage agreement of title or abstract screening was 97%. After screening, 51 full-text reports were assessed for eligibility, among which 45 reports were included in the systematic review [ 21 - 65 ]. The percentage agreement of the full-text review was 87%. In 40 of the included studies, contingency tables could not be made. Meta-analyses were conducted based on 8 AI models extracted from 6 studies. Individual study characteristics included in the systematic review are provided in Tables 1 and 2 . The facial feature extraction method can be categorized into 2 classes: geometrical features (GFs) and deep features (DFs). One typical method of extracting GFs is to calculate the distance between facial landmarks. DFs are usually extracted by convolution operations. A total of 20 studies included temporal information, but most of them (18) extracted temporal information through the 3D convolution of video sequences. Feature transformation was also commonly applied to reduce the time for training or fuse features extracted by different methods before inputting them into the classifier. For classifiers, support vector machines (SVMs) and convolutional neural networks (CNNs) were mostly used. Table 1 presents the model designs of the included studies.

a No temporal features are shown by – symbol, time information extracted from 2 images at different time by +, and deep temporal features extracted through the convolution of video sequences by ++.
b SVM: support vector machine.
c GF: geometric feature.
d GMM: gaussian mixture model.
e TPS: thin plate spline.
f DML: distance metric learning.
g MDML: multiview distance metric learning.
h AAM: active appearance model.
i RVR: relevance vector regressor.
j PSPI: Prkachin and Solomon pain intensity.
k I-FES: individual facial expressiveness score.
l LSTM: long short-term memory.
m HCRF: hidden conditional random field.
n GLMM: generalized linear mixed model.
o VLAD: vector of locally aggregated descriptor.
p SVR: support vector regression.
q MDS: multidimensional scaling.
r ELM: extreme learning machine.
s Labeled to distinguish different architectures of ensembled deep learning models.
t DCNN: deep convolutional neural network.
u GSM: gaussian scale mixture.
v DOML: distance ordering metric learning.
w LIAN: locality and identity aware network.
x BiLSTM: bidirectional long short-term memory.
a UNBC: University of Northern British Columbia-McMaster shoulder pain expression archive database.
b LOSO: leave one subject out cross-validation.
c ICC: intraclass correlation coefficient.
d CT: contingency table.
e AUC: area under the curve.
f MSE: mean standard error.
g PCC: Pearson correlation coefficient.
h RMSE: root mean standard error.
i MAE: mean absolute error.
j ICC: intraclass coefficient.
k CCC: concordance correlation coefficient.
l Reported both external and internal validation results and summarized as intervals.
Table 2 summarizes the characteristics of model training and validation. Most studies used publicly available databases, for example, the University of Northern British Columbia-McMaster shoulder pain expression archive database [ 57 ]. Table S4 in Multimedia Appendix 1 summarizes the public databases. A total of 7 studies used self-prepared databases. Frames from video sequences were the most used test objects, as 37 studies output frame-level pain intensity, while few measure pain intensity from video sequences or photos. It was common that a study redefined pain levels to have fewer classes than ground-truth labels. For model validation, cross-validation and leave-one-subject-out validation were commonly used. Only 3 studies performed external validation. For reporting test accuracies, different evaluation metrics were used, including sensitivity, specificity, mean absolute error (MAE), mean standard error (MSE), Pearson correlation coefficient (PCC), and intraclass coefficient (ICC).
Methodological Quality of Included Studies
Table S2 in Multimedia Appendix 1 presents the study quality summary, as assessed by QUADAS-2. There was a risk of bias in all studies, specifically in terms of patient selection, caused by 2 issues. First, the training data are highly imbalanced, and any method to adjust the data distribution may introduce bias. Next, the QUADAS-AI correspondence letter [ 19 ] specifies that preprocessing of images that changes the image size or resolution may introduce bias. However, the applicability concern is low, as the images properly represent the feeling of pain. Studies that used cross-fold validation or leave-one-out cross-validation were considered to have a low risk of bias. Although the Prkachin and Solomon pain intensity (PSPI) score was used by most of the studies, its ability to represent individual pain levels was not clinically validated; as such, the risk of bias and applicability concerns were considered high when the PSPI score was used as the index test. As an advantage of computer vision techniques, the time interval between the index tests was short and was assessed as having a low risk of bias. Risk proportions are shown in Figure 2 . For all 315 entries, 39% (124) were assessed as high-risk. In total, 5 studies had the lowest risk of bias, with 6 domains assessed as low risk [ 26 , 27 , 31 , 32 , 59 ].

Pooled Performance of Included Models
In 6 studies included in the meta-analysis, there were 8 different models. The characteristics of these models are summarized in Table S1 in Multimedia Appendix 2 [ 23 , 24 , 26 , 32 , 41 , 57 ]. Classification of PSPI scores greater than 0, 2, 3, 6, and 9 was selected and considered as different tasks to create contingency tables. The test performance is shown in Figure 3 as hierarchical SROC curves; 27 contingency tables were extracted from 8 models. The sensitivity, specificity, and LDOR were calculated, and the combined sensitivity was 98% (95% CI 96%-99%), the specificity was 98% (95% CI 97%-99%), the LDOR was 7.99 (95% CI 6.73-9.31) and the AUC was 0.99 (95% CI 0.99-1).

Subgroup Analysis
In this study, subgroup analysis was conducted to investigate the performance differences within models. A total of 8 models were separated and summarized as a forest plot in Multimedia Appendix 3 [ 23 , 24 , 26 , 32 , 41 , 57 ]. For model 1, the pooled sensitivity, specificity, and LDOR were 95% (95% CI 86%-99%), 99% (95% CI 98%-100%), and 8.38 (95% CI 6.09-11.19), respectively. For model 2, the pooled sensitivity, specificity, and LDOR were 94% (95% CI 84%-99%), 95% (95% CI 88%-99%), and 6.23 (95% CI 3.52-9.04), respectively. For model 3, the pooled sensitivity, specificity, and LDOR were 100% (95% CI 99%-100%), 100% (95% CI 99%-100%), and 11.55% (95% CI 8.82-14.43), respectively. For model 4, the pooled sensitivity, specificity, and LDOR were 83% (95% CI 43%-99%), 94% (95% CI 79%-99%), and 5.14 (95% CI 0.93-9.31), respectively. For model 5, the pooled sensitivity, specificity, and LDOR were 92% (95% CI 68%-99%), 94% (95% CI 78%-99%), and 6.12 (95% CI 1.82-10.16), respectively. For model 6, the pooled sensitivity, specificity, and LDOR were 94% (95% CI 74%-100%), 94% (95% CI 78%-99%), and 6.59 (95% CI 2.21-11.13), respectively. For model 7, the pooled sensitivity, specificity, and LDOR were 98% (95% CI 90%-100%), 97% (95% CI 87%-100%), and 8.31 (95% CI 4.3-12.29), respectively. For model 8, the pooled sensitivity, specificity, and LDOR were 98% (95% CI 93%-100%), 97% (95% CI 88%-100%), and 8.65 (95% CI 4.84-12.67), respectively.
Heterogeneity Analysis
The meta-analysis results indicated that AI models are applicable for estimating pain intensity from facial images. However, extreme heterogeneity existed within the models except for models 3 and 5, which were proposed by Rathee and Ganotra [ 24 ] and Semwal and Londhe [ 32 ]. A funnel plot is presented in Figure 4 . A high risk of bias was observed.

Pain management has long been a critical problem in clinical practice, and the use of AI may be a solution. For acute pain management, automatic measurement of pain can reduce the burden on caregivers and provide timely warnings. For chronic pain management, as specified by Glare et al [ 2 ], further research is needed, and measurements of pain presence, intensity, and quality are one of the issues to be solved for chronic pain studies. Computer vision could improve pain monitoring through real-time detection for clinical use and data recording for prospective pain studies. To our knowledge, this is the first meta-analysis dedicated to AI performance in multilevel pain level classification.
In this study, one model’s performance at specific pain levels was described by stacking multiple classes into one to make each task a binary classification problem. After careful selection in both the medical and engineering databases, we observed promising results of AI in evaluating multilevel pain intensity through facial images, with high sensitivity (98%), specificity (98%), LDOR (7.99), and AUC (0.99). It is reasonable to believe that AI can accurately evaluate pain intensity from facial images. Moreover, the study quality and risk of bias were evaluated using an adapted QUADAS-2 assessment tool, which is a strength of this study.
To investigate the source of heterogeneity, it was assumed that a well-designed model should have familiar size effects regarding different levels, and a subgroup meta-analysis was conducted. The funnel and forest plots exhibited extreme heterogeneity. The model’s performance at specific pain levels was described and summarized by a forest plot. Within-model heterogeneity was observed in Multimedia Appendix 3 [ 23 , 24 , 26 , 32 , 41 , 57 ] except for 2 models. Models 3 and 5 were different in many aspects, including their algorithms and validation methods, but were both trained with a relatively small data set, and the proportion of positive and negative classes was relatively close to 1. Because training with imbalanced data is a critical problem in computer vision studies [ 66 ], for example, in the University of Northern British Columbia-McMaster pain data set, fewer than 10 frames out of 48,398 had a PSPI score greater than 13. Here, we emphasized that imbalanced data sets are one major cause of heterogeneity, resulting in the poorer performance of AI algorithms.
We tentatively propose a method to minimize the effect of training with imbalanced data by stacking multiple classes into one class, which is already presented in studies included in the systematic review [ 26 , 32 , 42 , 57 ]. Common methods to minimize bias include resampling and data augmentation [ 66 ]. This proposed method is used in the meta-analysis to compare the test results of different studies as well. The stacking method is available when classes are only different in intensity. A disadvantage of combined classes is that the model would be insufficient in clinical practice when the number of classes is low. Commonly used pain evaluation tools, such as VAS, have 10 discrete levels. It is recommended that future studies set the number of pain levels to be at least 10 for model training.
This study is limited for several reasons. First, insufficient data were included because different performance metrics (mean standard error and mean average error) were used in most studies, which could not be summarized into a contingency table. To create a contingency table that can be included in a meta-analysis, the study should report the following: the number of objects used in each pain class for model validation, and the accuracy, sensitivity, specificity, and F 1 -score for each pain class. This table cannot be created if a study reports the MAE, PCC, and other commonly used metrics in AI development. Second, a small study effect was observed in the funnel plot, and the heterogeneity could not be minimized. Another limitation is that the PSPI score is not clinically validated and is not the only tool that assesses pain from facial expressions. There are other clinically validated pain intensity assessment methods, such as the Faces Pain Scale-revised, Wong-Baker Faces Pain Rating Scale, and Oucher Scale [ 3 ]. More databases could be created based on the above-mentioned tools. Finally, AI-assisted pain assessments were supposed to cover larger populations, including incommunicable patients, for example, patients with dementia or patients with masked faces. However, only 1 study considered patients with dementia, which was also caused by limited databases [ 50 ].
AI is a promising tool that can help in pain research in the future. In this systematic review and meta-analysis, one approach using computer vision was investigated to measure pain intensity from facial images. Despite some risk of bias and applicability concerns, CV models can achieve excellent test accuracy. Finally, more CV studies in pain estimation, reporting accuracy in contingency tables, and more pain databases are encouraged for future studies. Specifically, the creation of a balanced public database that contains not only healthy but also nonhealthy participants should be prioritized. The recording process would be better in a clinical environment. Then, it is recommended that researchers report the validation results in terms of accuracy, sensitivity, specificity, or contingency tables, as well as the number of objects for each pain class, for the inclusion of a meta-analysis.
Acknowledgments
WL, AH, and CW contributed to the literature search and data extraction. JH and YY wrote the first draft of the manuscript. All authors contributed to the conception and design of the study, the risk of bias evaluation, data analysis and interpretation, and contributed to and approved the final version of the manuscript.
Data Availability
The data sets generated during and analyzed during this study are available in the Figshare repository [ 67 ].
Conflicts of Interest
None declared.
PRISMA checklist, risk of bias summary, search strategy, database summary and reported items and explanations.
Study performance summary.
Forest plot presenting pooled performance of subgroups in meta-analysis.
- Raja SN, Carr DB, Cohen M, Finnerup NB, Flor H, Gibson S, et al. The revised International Association for the Study of Pain definition of pain: concepts, challenges, and compromises. Pain. 2020;161(9):1976-1982. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Glare P, Aubrey KR, Myles PS. Transition from acute to chronic pain after surgery. Lancet. 2019;393(10180):1537-1546. [ CrossRef ] [ Medline ]
- Chou R, Gordon DB, de Leon-Casasola OA, Rosenberg JM, Bickler S, Brennan T, et al. Management of postoperative pain: a clinical practice guideline from the American Pain Society, the American Society of Regional Anesthesia and Pain Medicine, and the American Society of Anesthesiologists' Committee on Regional Anesthesia, Executive Committee, and Administrative Council. J Pain. 2016;17(2):131-157. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Hassan T, Seus D, Wollenberg J, Weitz K, Kunz M, Lautenbacher S, et al. Automatic detection of pain from facial expressions: a survey. IEEE Trans Pattern Anal Mach Intell. 2021;43(6):1815-1831. [ CrossRef ] [ Medline ]
- Mussigmann T, Bardel B, Lefaucheur JP. Resting-State Electroencephalography (EEG) biomarkers of chronic neuropathic pain. A systematic review. Neuroimage. 2022;258:119351. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Moscato S, Cortelli P, Chiari L. Physiological responses to pain in cancer patients: a systematic review. Comput Methods Programs Biomed. 2022;217:106682. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Thiam P, Hihn H, Braun DA, Kestler HA, Schwenker F. Multi-modal pain intensity assessment based on physiological signals: a deep learning perspective. Front Physiol. 2021;12:720464. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Rojas RF, Brown N, Waddington G, Goecke R. A systematic review of neurophysiological sensing for the assessment of acute pain. NPJ Digit Med. 2023;6(1):76. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Mansutti I, Tomé-Pires C, Chiappinotto S, Palese A. Facilitating pain assessment and communication in people with deafness: a systematic review. BMC Public Health. 2023;23(1):1594. [ FREE Full text ] [ CrossRef ] [ Medline ]
- El-Tallawy SN, Ahmed RS, Nagiub MS. Pain management in the most vulnerable intellectual disability: a review. Pain Ther. 2023;12(4):939-961. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Gkikas S, Tsiknakis M. Automatic assessment of pain based on deep learning methods: a systematic review. Comput Methods Programs Biomed. 2023;231:107365. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Borna S, Haider CR, Maita KC, Torres RA, Avila FR, Garcia JP, et al. A review of voice-based pain detection in adults using artificial intelligence. Bioengineering (Basel). 2023;10(4):500. [ FREE Full text ] [ CrossRef ] [ Medline ]
- De Sario GD, Haider CR, Maita KC, Torres-Guzman RA, Emam OS, Avila FR, et al. Using AI to detect pain through facial expressions: a review. Bioengineering (Basel). 2023;10(5):548. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Zhang M, Zhu L, Lin SY, Herr K, Chi CL, Demir I, et al. Using artificial intelligence to improve pain assessment and pain management: a scoping review. J Am Med Inform Assoc. 2023;30(3):570-587. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Hughes JD, Chivers P, Hoti K. The clinical suitability of an artificial intelligence-enabled pain assessment tool for use in infants: feasibility and usability evaluation study. J Med Internet Res. 2023;25:e41992. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Fang J, Wu W, Liu J, Zhang S. Deep learning-guided postoperative pain assessment in children. Pain. 2023;164(9):2029-2035. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Page MJ, McKenzie JE, Bossuyt PM, Boutron I, Hoffmann TC, Mulrow CD, et al. The PRISMA 2020 statement: an updated guideline for reporting systematic reviews. BMJ. 2021;372:n71. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Whiting PF, Rutjes AWS, Westwood ME, Mallett S, Deeks JJ, Reitsma JB, et al. QUADAS-2: a revised tool for the quality assessment of diagnostic accuracy studies. Ann Intern Med. 2011;155(8):529-536. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Sounderajah V, Ashrafian H, Rose S, Shah NH, Ghassemi M, Golub R, et al. A quality assessment tool for artificial intelligence-centered diagnostic test accuracy studies: QUADAS-AI. Nat Med. 2021;27(10):1663-1665. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Guo J, Riebler A. meta4diag: Bayesian bivariate meta-analysis of diagnostic test studies for routine practice. J Stat Soft. 2018;83(1):1-31. [ CrossRef ]
- Hammal Z, Cohn JF. Automatic detection of pain intensity. Proc ACM Int Conf Multimodal Interact. 2012;2012:47-52. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Adibuzzaman M, Ostberg C, Ahamed S, Povinelli R, Sindhu B, Love R, et al. Assessment of pain using facial pictures taken with a smartphone. 2015. Presented at: 2015 IEEE 39th Annual Computer Software and Applications Conference; July 01-05, 2015;726-731; Taichung, Taiwan. [ CrossRef ]
- Majumder A, Dutta S, Behera L, Subramanian VK. Shoulder pain intensity recognition using Gaussian mixture models. 2015. Presented at: 2015 IEEE International WIE Conference on Electrical and Computer Engineering (WIECON-ECE); December 19-20, 2015;130-134; Dhaka, Bangladesh. [ CrossRef ]
- Rathee N, Ganotra D. A novel approach for pain intensity detection based on facial feature deformations. J Vis Commun Image Represent. 2015;33:247-254. [ CrossRef ]
- Sikka K, Ahmed AA, Diaz D, Goodwin MS, Craig KD, Bartlett MS, et al. Automated assessment of children's postoperative pain using computer vision. Pediatrics. 2015;136(1):e124-e131. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Rathee N, Ganotra D. Multiview distance metric learning on facial feature descriptors for automatic pain intensity detection. Comput Vis Image Und. 2016;147:77-86. [ CrossRef ]
- Zhou J, Hong X, Su F, Zhao G. Recurrent convolutional neural network regression for continuous pain intensity estimation in video. 2016. Presented at: 2016 IEEE Conference on Computer Vision and Pattern Recognition Workshops (CVPRW); June 26-July 01, 2016; Las Vegas, NV. [ CrossRef ]
- Egede J, Valstar M, Martinez B. Fusing deep learned and hand-crafted features of appearance, shape, and dynamics for automatic pain estimation. 2017. Presented at: 2017 12th IEEE International Conference on Automatic Face & Gesture Recognition (FG 2017); May 30-June 03, 2017;689-696; Washington, DC. [ CrossRef ]
- Martinez DL, Rudovic O, Picard R. Personalized automatic estimation of self-reported pain intensity from facial expressions. 2017. Presented at: 2017 IEEE Conference on Computer Vision and Pattern Recognition Workshops (CVPRW); July 21-26, 2017;2318-2327; Honolulu, HI. [ CrossRef ]
- Bourou D, Pampouchidou A, Tsiknakis M, Marias K, Simos P. Video-based pain level assessment: feature selection and inter-subject variability modeling. 2018. Presented at: 2018 41st International Conference on Telecommunications and Signal Processing (TSP); July 04-06, 2018;1-6; Athens, Greece. [ CrossRef ]
- Haque MA, Bautista RB, Noroozi F, Kulkarni K, Laursen C, Irani R. Deep multimodal pain recognition: a database and comparison of spatio-temporal visual modalities. 2018. Presented at: 2018 13th IEEE International Conference on Automatic Face & Gesture Recognition (FG 2018); May 15-19, 2018;250-257; Xi'an, China. [ CrossRef ]
- Semwal A, Londhe ND. Automated pain severity detection using convolutional neural network. 2018. Presented at: 2018 International Conference on Computational Techniques, Electronics and Mechanical Systems (CTEMS); December 21-22, 2018;66-70; Belgaum, India. [ CrossRef ]
- Tavakolian M, Hadid A. Deep binary representation of facial expressions: a novel framework for automatic pain intensity recognition. 2018. Presented at: 2018 25th IEEE International Conference on Image Processing (ICIP); October 07-10, 2018;1952-1956; Athens, Greece. [ CrossRef ]
- Tavakolian M, Hadid A. Deep spatiotemporal representation of the face for automatic pain intensity estimation. 2018. Presented at: 2018 24th International Conference on Pattern Recognition (ICPR); August 20-24, 2018;350-354; Beijing, China. [ CrossRef ]
- Wang J, Sun H. Pain intensity estimation using deep spatiotemporal and handcrafted features. IEICE Trans Inf & Syst. 2018;E101.D(6):1572-1580. [ CrossRef ]
- Bargshady G, Soar J, Zhou X, Deo RC, Whittaker F, Wang H. A joint deep neural network model for pain recognition from face. 2019. Presented at: 2019 IEEE 4th International Conference on Computer and Communication Systems (ICCCS); February 23-25, 2019;52-56; Singapore. [ CrossRef ]
- Casti P, Mencattini A, Comes MC, Callari G, Di Giuseppe D, Natoli S, et al. Calibration of vision-based measurement of pain intensity with multiple expert observers. IEEE Trans Instrum Meas. 2019;68(7):2442-2450. [ CrossRef ]
- Lee JS, Wang CW. Facial pain intensity estimation for ICU patient with partial occlusion coming from treatment. 2019. Presented at: BIBE 2019; The Third International Conference on Biological Information and Biomedical Engineering; June 20-22, 2019;1-4; Hangzhou, China.
- Saha AK, Ahsan GMT, Gani MO, Ahamed SI. Personalized pain study platform using evidence-based continuous learning tool. 2019. Presented at: 2019 IEEE 43rd Annual Computer Software and Applications Conference (COMPSAC); July 15-19, 2019;490-495; Milwaukee, WI. [ CrossRef ]
- Tavakolian M, Hadid A. A spatiotemporal convolutional neural network for automatic pain intensity estimation from facial dynamics. Int J Comput Vis. 2019;127(10):1413-1425. [ FREE Full text ] [ CrossRef ]
- Bargshady G, Zhou X, Deo RC, Soar J, Whittaker F, Wang H. Ensemble neural network approach detecting pain intensity from facial expressions. Artif Intell Med. 2020;109:101954. [ CrossRef ] [ Medline ]
- Bargshady G, Zhou X, Deo RC, Soar J, Whittaker F, Wang H. Enhanced deep learning algorithm development to detect pain intensity from facial expression images. Expert Syst Appl. 2020;149:113305. [ CrossRef ]
- Dragomir MC, Florea C, Pupezescu V. Automatic subject independent pain intensity estimation using a deep learning approach. 2020. Presented at: 2020 International Conference on e-Health and Bioengineering (EHB); October 29-30, 2020;1-4; Iasi, Romania. [ CrossRef ]
- Huang D, Xia Z, Mwesigye J, Feng X. Pain-attentive network: a deep spatio-temporal attention model for pain estimation. Multimed Tools Appl. 2020;79(37-38):28329-28354. [ CrossRef ]
- Mallol-Ragolta A, Liu S, Cummins N, Schuller B. A curriculum learning approach for pain intensity recognition from facial expressions. 2020. Presented at: 2020 15th IEEE International Conference on Automatic Face and Gesture Recognition (FG 2020); November 16-20, 2020;829-833; Buenos Aires, Argentina. [ CrossRef ]
- Peng X, Huang D, Zhang H. Pain intensity recognition via multi‐scale deep network. IET Image Process. 2020;14(8):1645-1652. [ FREE Full text ] [ CrossRef ]
- Tavakolian M, Lopez MB, Liu L. Self-supervised pain intensity estimation from facial videos via statistical spatiotemporal distillation. Pattern Recognit Lett. 2020;140:26-33. [ CrossRef ]
- Xu X, de Sa VR. Exploring multidimensional measurements for pain evaluation using facial action units. 2020. Presented at: 2020 15th IEEE International Conference on Automatic Face and Gesture Recognition (FG 2020); November 16-20, 2020;786-792; Buenos Aires, Argentina. [ CrossRef ]
- Pikulkaew K, Boonchieng W, Boonchieng E, Chouvatut V. 2D facial expression and movement of motion for pain identification with deep learning methods. IEEE Access. 2021;9:109903-109914. [ CrossRef ]
- Rezaei S, Moturu A, Zhao S, Prkachin KM, Hadjistavropoulos T, Taati B. Unobtrusive pain monitoring in older adults with dementia using pairwise and contrastive training. IEEE J Biomed Health Inform. 2021;25(5):1450-1462. [ CrossRef ] [ Medline ]
- Semwal A, Londhe ND. S-PANET: a shallow convolutional neural network for pain severity assessment in uncontrolled environment. 2021. Presented at: 2021 IEEE 11th Annual Computing and Communication Workshop and Conference (CCWC); January 27-30, 2021;0800-0806; Las Vegas, NV. [ CrossRef ]
- Semwal A, Londhe ND. ECCNet: an ensemble of compact convolution neural network for pain severity assessment from face images. 2021. Presented at: 2021 11th International Conference on Cloud Computing, Data Science & Engineering (Confluence); January 28-29, 2021;761-766; Noida, India. [ CrossRef ]
- Szczapa B, Daoudi M, Berretti S, Pala P, Del Bimbo A, Hammal Z. Automatic estimation of self-reported pain by interpretable representations of motion dynamics. 2021. Presented at: 2020 25th International Conference on Pattern Recognition (ICPR); January 10-15, 2021;2544-2550; Milan, Italy. [ CrossRef ]
- Ting J, Yang YC, Fu LC, Tsai CL, Huang CH. Distance ordering: a deep supervised metric learning for pain intensity estimation. 2021. Presented at: 2021 20th IEEE International Conference on Machine Learning and Applications (ICMLA); December 13-16, 2021;1083-1088; Pasadena, CA. [ CrossRef ]
- Xin X, Li X, Yang S, Lin X, Zheng X. Pain expression assessment based on a locality and identity aware network. IET Image Process. 2021;15(12):2948-2958. [ FREE Full text ] [ CrossRef ]
- Alghamdi T, Alaghband G. Facial expressions based automatic pain assessment system. Appl Sci. 2022;12(13):6423. [ FREE Full text ] [ CrossRef ]
- Barua PD, Baygin N, Dogan S, Baygin M, Arunkumar N, Fujita H, et al. Automated detection of pain levels using deep feature extraction from shutter blinds-based dynamic-sized horizontal patches with facial images. Sci Rep. 2022;12(1):17297. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Fontaine D, Vielzeuf V, Genestier P, Limeux P, Santucci-Sivilotto S, Mory E, et al. Artificial intelligence to evaluate postoperative pain based on facial expression recognition. Eur J Pain. 2022;26(6):1282-1291. [ CrossRef ] [ Medline ]
- Hosseini E, Fang R, Zhang R, Chuah CN, Orooji M, Rafatirad S, et al. Convolution neural network for pain intensity assessment from facial expression. 2022. Presented at: 2022 44th Annual International Conference of the IEEE Engineering in Medicine & Biology Society (EMBC); July 11-15, 2022;2697-2702; Glasgow, Scotland. [ CrossRef ]
- Huang Y, Qing L, Xu S, Wang L, Peng Y. HybNet: a hybrid network structure for pain intensity estimation. Vis Comput. 2021;38(3):871-882. [ CrossRef ]
- Islamadina R, Saddami K, Oktiana M, Abidin TF, Muharar R, Arnia F. Performance of deep learning benchmark models on thermal imagery of pain through facial expressions. 2022. Presented at: 2022 IEEE International Conference on Communication, Networks and Satellite (COMNETSAT); November 03-05, 2022;374-379; Solo, Indonesia. [ CrossRef ]
- Swetha L, Praiscia A, Juliet S. Pain assessment model using facial recognition. 2022. Presented at: 2022 6th International Conference on Intelligent Computing and Control Systems (ICICCS); May 25-27, 2022;1-5; Madurai, India. [ CrossRef ]
- Wu CL, Liu SF, Yu TL, Shih SJ, Chang CH, Mao SFY, et al. Deep learning-based pain classifier based on the facial expression in critically ill patients. Front Med (Lausanne). 2022;9:851690. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Ismail L, Waseem MD. Towards a deep learning pain-level detection deployment at UAE for patient-centric-pain management and diagnosis support: framework and performance evaluation. Procedia Comput Sci. 2023;220:339-347. [ FREE Full text ] [ CrossRef ] [ Medline ]
- Vu MT, Beurton-Aimar M. Learning to focus on region-of-interests for pain intensity estimation. 2023. Presented at: 2023 IEEE 17th International Conference on Automatic Face and Gesture Recognition (FG); January 05-08, 2023;1-6; Waikoloa Beach, HI. [ CrossRef ]
- Kaur H, Pannu HS, Malhi AK. A systematic review on imbalanced data challenges in machine learning: applications and solutions. ACM Comput Surv. 2019;52(4):1-36. [ CrossRef ]
- Data for meta-analysis of pain assessment from facial images. Figshare. 2023. URL: https://figshare.com/articles/dataset/Data_for_Meta-Analysis_of_Pain_Assessment_from_Facial_Images/24531466/1 [accessed 2024-03-22]
Abbreviations
Edited by A Mavragani; submitted 26.07.23; peer-reviewed by M Arab-Zozani, M Zhang; comments to author 18.09.23; revised version received 08.10.23; accepted 28.02.24; published 12.04.24.
©Jian Huo, Yan Yu, Wei Lin, Anmin Hu, Chaoran Wu. Originally published in the Journal of Medical Internet Research (https://www.jmir.org), 12.04.2024.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work, first published in the Journal of Medical Internet Research, is properly cited. The complete bibliographic information, a link to the original publication on https://www.jmir.org/, as well as this copyright and license information must be included.
IMAGES
VIDEO
COMMENTS
Biodiversity is the variety of different forms of life on earth, including the different plants, animals, micro-organisms, the. genes they contain and the ecosystem they form. It refers to genetic ...
Towards a Biodiversity Conservation Research Strategy. Over 40 research topics are identified in the chapters of this volume. Such compilations of research opportunities need to be embraced within national strategies, matching needs with resources and priorities within a research agenda, while understanding the country's challenges.
Biodiversity is a crucial part of nature's precious assets that provide many human needs and insures against environmental disasters. Scientists have not yet reached a consensus on the definition ...
1. Introduction. While conservation biologists have been concerned with the impacts of human activities on biodiversity since decades (Soulé, 1985), biodiversity loss is increasingly being recognized as one of the top risks that humanity is facing by a variety of actors (United Nations, 2020; Robuchon et al., 2021; WEF, 2023).According to recent major global biodiversity assessments (IPBES ...
Conclusion: biodiversity requires diversity. ... where a first draft of this paper was presented and for editing the special issue of which this paper is part. ... 'Satellite Remote Sensing, Biodiversity Research and Conservation of the Future' 369 Philosophical Transactions of the Royal Society B: Biological Sciences 20130190.
Conclusion: As biodiversity underlies all goods and services provided by ecosystems that are crucial for human survival and well-being, this paper synthesizes and discusses observed and anticipated impacts of climate change on ... C., C. Folke, and K. G. Mäler. 1992. "The Ecology and Economics of Biodiversity Loss: The Research Agenda." ...
Categorization of papers to address questions 2 and 3 was determined primarily from our expert judgement, as few papers were explicit about the scales at which biodiversity benefits are delivered.
Concerns over the consequences of biodiversity loss for human well-being triggered the growth of biodiversity-ecosystem functioning (BEF) research, an important field of ecology over the past 25 ...
Recent global reports (Díaz et al. 2019; IPBES Secretariat 2019; CBD 2020) have rigorously synthesized the large scientific literature on biodiversity and have identified major knowledge gaps.These gaps include large uncertainties in how many species are threatened with extinction (Díaz et al. 2019; CBD 2020; IUCN 2020), a lack of estimates for the impacts of global biodiversity loss on ...
The social, economic, and environmental implications of biodiversity change have been the focus of interdisciplinary research for the last three decades 4,5. For example, influential experiments have revealed that species loss can reduce the productivity and stability of plant communities. Similarly, declines in biological diversity can also ...
Conservation biology is designed to identify pressing environmental problems and to solve them. This review evaluates the relative effort of conservation biology in problem-based and solution-based research, and tests whether or not this has changed in the past decades for five major drivers of biodiversity loss, i.e. habitat loss and fragmentation, overexploitation, biological invasion ...
1. Introduction. The need to feed, clothe and house humanity has altered most of the Earth's ecosystems [].The dominant anthropogenic processes include the conversion of natural landscapes for agriculture and urbanization, the accidental or purposeful introduction of non-native species, and direct harvesting of wild populations ().These activities have greatly altered the composition of biotic ...
Abstract. As society strives to transition towards more sustainable development pathways, it is important to properly conceptualize the link between biodiversity (i.e. genes, traits, species and other dimensions) and human well-being (HWB; i.e. health, wealth, security and other dimensions). Here, we explore how published conceptual frameworks ...
Introduction. Habitat loss has been, and still is, the greatest threat to biodiversity (Brooks et al. 2002; Hanski 2005; Groom et al. 2006).According to the Millennium Ecosystem Assessment (), more than half of several biomes, including the Mediterranean and temperate forests and tropical and sub-tropical dry broadleaf forests, had been converted by 1990; in Western Europe, only 2-3% of ...
The lack of research effort in key areas of high priority in the threats to soil biodiversity are a concerning finding and requires some consideration and debate in the research community. ... 46 papers were left for review. Each reference was studied in detail, and the potential threats to soil biodiversity that each paper identified was ...
1. Introduction. Climate change is a pervasive and growing global threat to biodiversity and ecosystems (Díaz et al., 2019).Climate change affects individual species and the way they interact with other organisms and their habitats, which alters the structure and function of ecosystems and the goods and services that natural systems provide to society (Díaz et al., 2019).
Over the last 25 years, research on biodiversity has expanded dramatically, fuelled by increasing threats to the natural world. However, the number of published studies is heavily weighted towards certain taxa, perhaps influencing conservation awareness of and funding for less-popular groups. Few studies have systematically quantified these biases, although information on this topic is ...
Abstract: Biodiversity is the amount of variety of life on Earth. It is the number of different species of plants, animals, and microorganisms surviving on our planet. It includes organisms from ...
Go-Away-Birding, Cape Town 20 Conclusions: Biodiversity Research and Conservation Opportunities 549 Vaz da Silva B (2015) Evolutionary history of the birds of the Angolan highlands - the missing piece to understand the biogeography of the Afromontane forests. MSc thesis.
INTRODUCTION. One of the greatest attributes of the Earth is the biodiversity of her ecosystem. The Convention on Biological Diversity (Article 2) defined biological diversity or biodiversity" as the variability among living organisms from all sources including, inter alia, terrestrial, marine and other aquatic ecosystems and the ecological complexes of which they are part; this includes ...
Abstract. Human beings are not only a part of our planet's ecosystems, but also, they are massively overusing them. This makes ecosystem protection, including biodiversity preservation, vital for humanity's future. The speed and scale of the threat are unprecedented in human history. The long arch of evolution has been confronted with such a ...
This sample diversity research paper on biodiversity features: 4800 words (approx. 16 pages) and a bibliography with 12 sources. Browse other research paper examples for more inspiration. ... Conclusion. Considering things at the planetary scale, it is now assessed that biodiversity losses and its corollary, the deterioration of ecosystem ...
The field of biodiversity studies now embraces the activities of many disciplines, including paleobiology, systematics, ecology, and conservation biology. Special emphasis given on biodiversity ...
Background: The continuous monitoring and recording of patients' pain status is a major problem in current research on postoperative pain management. In the large number of original or review articles focusing on different approaches for pain assessment, many researchers have investigated how computer vision (CV) can help by capturing facial expressions.