
An official website of the United States government
The .gov means it’s official. Federal government websites often end in .gov or .mil. Before sharing sensitive information, make sure you’re on a federal government site.
The site is secure. The https:// ensures that you are connecting to the official website and that any information you provide is encrypted and transmitted securely.
- Publications
- Account settings
Preview improvements coming to the PMC website in October 2024. Learn More or Try it out now .
- Advanced Search
- Journal List
- Front Hum Neurosci
- PMC10410470

Cognitive neuroscience perspective on memory: overview and summary
Sruthi sridhar.
1 Department of Psychology, Mount Allison University, Sackville, NB, Canada
Abdulrahman Khamaj
2 Department of Industrial Engineering, College of Engineering, Jazan University, Jazan, Saudi Arabia
Manish Kumar Asthana
3 Department of Humanities and Social Sciences, Indian Institute of Technology Roorkee, Roorkee, India
4 Department of Design, Indian Institute of Technology Roorkee, Roorkee, India
Associated Data
The original contributions presented in this study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
This paper explores memory from a cognitive neuroscience perspective and examines associated neural mechanisms. It examines the different types of memory: working, declarative, and non-declarative, and the brain regions involved in each type. The paper highlights the role of different brain regions, such as the prefrontal cortex in working memory and the hippocampus in declarative memory. The paper also examines the mechanisms that underlie the formation and consolidation of memory, including the importance of sleep in the consolidation of memory and the role of the hippocampus in linking new memories to existing cognitive schemata. The paper highlights two types of memory consolidation processes: cellular consolidation and system consolidation. Cellular consolidation is the process of stabilizing information by strengthening synaptic connections. System consolidation models suggest that memories are initially stored in the hippocampus and are gradually consolidated into the neocortex over time. The consolidation process involves a hippocampal-neocortical binding process incorporating newly acquired information into existing cognitive schemata. The paper highlights the role of the medial temporal lobe and its involvement in autobiographical memory. Further, the paper discusses the relationship between episodic and semantic memory and the role of the hippocampus. Finally, the paper underscores the need for further research into the neurobiological mechanisms underlying non-declarative memory, particularly conditioning. Overall, the paper provides a comprehensive overview from a cognitive neuroscience perspective of the different processes involved in memory consolidation of different types of memory.
Introduction
Memory is an essential cognitive function that permits individuals to acquire, retain, and recover data that defines a person’s identity ( Zlotnik and Vansintjan, 2019 ). Memory is a multifaceted cognitive process that involves different stages: encoding, consolidation, recovery, and reconsolidation. Encoding involves acquiring and processing information that is transformed into a neuronal representation suitable for storage ( Liu et al., 2021 ; Panzeri et al., 2023 ). The information can be acquired through various channels, such as visual, auditory, olfactory, or tactile inputs. The acquired sensory stimuli are converted into a format the brain can process and retain. Different factors such as attention, emotional significance, and repetition can influence the encoding process and determine the strength and durability of the resulting memory ( Squire et al., 2004 ; Lee et al., 2016 ; Serences, 2016 ).
Consolidation includes the stabilization and integration of memory into long-term storage to increase resistance to interference and decay ( Goedert and Willingham, 2002 ). This process creates enduring structural modification in the brain and thereby has consequential effects on the function by reorganizing and strengthening neural connections. Diverse sources like sleep and stress and the release of neurotransmitters can influence memory consolidation. Many researchers have noted the importance of sleep due to its critical role in enabling a smooth transition of information from transient repositories into more stable engrams (memory traces) ( McGaugh, 2000 ; Clawson et al., 2021 ; Rakowska et al., 2022 ).
Retrieval involves accessing, selecting, and reactivating or reconstructing the stored memory to allow conscious access to previously encoded information ( Dudai, 2002 ). Retrieving memories depends on activating relevant neural pathways while reconstructing encoded information. Factors like contextual or retrieval cues and familiarity with the material can affect this process. Forgetting becomes a possibility if there are inadequate triggers for associated memory traces to activate upon recall. Luckily, mnemonic strategies and retrieval practice offer effective tools to enhance recovery rates and benefit overall memory performance ( Roediger and Butler, 2011 ).
Previous research implied that once a memory has been consolidated, it becomes permanent ( McGaugh, 2000 ; Robins, 2020 ). However, recent studies have found an additional phase called “reconsolidation,” during which stored memories, when reactivated, enter a fragile or liable state and become susceptible to modification or update ( Schiller et al., 2009 ; Asthana et al., 2015 ). The process highlights the notion that memory is not static but a dynamic system influenced by subsequent encounters. The concept of reconsolidation has much significance in memory modification therapies and interventions, as it offers a promising opportunity to target maladaptive or traumatic memories for modification specifically. However, more thorough investigations are needed to gain insight into the mechanisms and concrete implications of employing memory reconsolidation within therapeutic settings ( Bellfy and Kwapis, 2020 ).
The concept of memory is not reducible to a single unitary phenomenon; instead, evidence suggests that it can be subdivided into several distinct but interrelated constituent processes and systems ( Richter-Levin and Akirav, 2003 ). There are three major types of human memory: working memory, declarative memory (explicit), and non-declarative memory (implicit). All these types of memories involve different neural systems in the brain. Working memory is a unique transient active store capable of manipulating information essential for many complex cognitive operations, including language processing, reasoning, and judgment ( Atkinson and Shiffrin, 1968 ; Baddeley and Logie, 1999 ; Funahashi, 2017 ; Quentin et al., 2019 ). Previous models suggest the existence of three components that make up the working memory ( Baddeley and Hitch, 1974 ; Baddeley, 1986 ). One master component, the central executive, controls the two dependent components, the phonological loop (speech perception and language comprehension) and the visuospatial sketchpad (visual images and spatial impressions processing). Some models mention a third component known as the episodic buffer. It is theorized that the episodic buffer serves as an intermediary between perception, long-term memory, and two components of working memory (the phonological loop and visuospatial sketchpad) by storing integrated episodes or chunks from both sources ( Baddeley, 2000 ). Declarative memory (explicit memory) can be recalled consciously, including facts and events that took place in one’s life or information learned from books. It encompasses memories of both autobiographical experiences and memories associated with general knowledge. It is usually associated with the hippocampus–medial temporal lobe system ( Thompson and Kim, 1996 ; Ober, 2014 ). Non-declarative memory (implicit memory) refers to unconscious forms of learning such as skills, habits, and priming effects; this type of implicit learning does not involve conscious recollection but can include motor skill tasks that often require no thought prior to execution nor later recall upon completion. This type of memory usually involves the amygdala and other systems ( Thompson and Kim, 1996 ; Ober, 2014 ).
Working memory
Working memory is primarily associated with the prefrontal and posterior parietal cortex ( Sarnthein et al., 1998 ; Todd and Marois, 2005 ). Working memory is not localized to a single brain region, and research suggests that it is an emergent property arising from functional interactions between the prefrontal cortex (PFC) and the rest of the brain ( D’Esposito, 2007 ). Neuroimaging studies have explored the neural basis for the three components proposed by Baddeley and Hitch (1974) , the Central executive, the phonological loop, and the visuospatial sketch pad; there is evidence for the existence of a fourth component called the episodic buffer ( Baddeley, 2000 ).
The central executive plays a significant role in working memory by acting as the control center ( Shallice, 2002 ). It facilitates critical functions like attention allocation and coordination between the phonological loop and the visuospatial sketchpad ( Yu et al., 2023 ). Recent findings have illuminated the dual-functional network regulation, the cingulo-opercular network (CON) and the frontoparietal network (FPN), that underpins the central executive system ( Yu et al., 2023 ). The CON comprises the dorsal anterior cingulate cortex (dACC) and anterior insula (AI). In contrast, the FPN encompasses various regions, such as the dorsolateral prefrontal cortex (DLPFC) and frontal eye field (FEF), along with the intraparietal sulcus (IPS) ( Yu et al., 2023 ). Neuroimaging research has found evidence that elucidates the neural underpinnings of the executive attention control system to the dorsolateral prefrontal cortex (DLPFC) and the anterior cingulate cortex (ACC) ( Jung et al., 2022 ). The activation patterns indicate that the CON may have a broader top-down control function across the working memory process. At the same time, the FPN could be more heavily implicated in momentary control or processing at the trial level ( Yu et al., 2023 ). Evidence suggests that the central executive interacts with the phonological loop and visuospatial sketchpad to support working memory processes ( Baddeley, 2003 ; Buchsbaum, 2010 ; Menon and D’Esposito, 2021 ). The function, localization, and neural basis of this interaction are thought to involve the activation of specific brain regions associated with each component of working memory, as discussed in detail below.
The phonological loop is divided into two components: a storage system that maintains information (a few seconds) and a component involving subvocal rehearsal—which maintains and refreshes information in the working memory. Neuroanatomically, the phonological loop is represented in the Brodmann area (BA) 40 in the parietal cortex and the rehearsal components in BA 44 and 6, both situated in the frontal cortex ( Osaka et al., 2007 ). The left inferior frontal gyrus (Broca’s area) and the left posterior superior temporal gyrus (Wernicke’s area) has been proposed to play a critical role in supporting phonological and verbal working memory tasks, specifically the subvocal rehearsal system of the articulatory loop ( Paulesu et al., 1993 ; Buchsbaum et al., 2001 ; Perrachione et al., 2017 ). The phonological store in verbal short-term memory has been localized at the left supramarginal gyrus ( Graves et al., 2008 ; Perrachione et al., 2017 ).
Studies utilizing neuroimaging techniques have consistently yielded results indicating notable activation in these brain regions during phonological activities like recalling non-words and maintaining verbal information in memory ( Awh et al., 1996 ; Graves et al., 2008 ). During tasks that require phonological rehearsal, there was an increase in activation in the left inferior frontal gyrus ( Paulesu et al., 1993 ). Researchers have noted an increase in activity within the superior temporal gyrus-which plays a significant role in auditory processing-in individuals performing tasks that necessitate verbal information maintenance and manipulation ( Smith et al., 1998 ; Chein et al., 2003 ).
Additionally, lesion studies have provided further confirmation regarding the importance of these regions. These investigations have revealed that impairment in performing phonological working memory tasks can transpire following damage inflicted upon the left hemisphere, particularly on perisylvian language areas ( Koenigs et al., 2011 ). It is common for individuals with lesions affecting regions associated with the phonological loop, such as the left inferior frontal gyrus and superior temporal gyrus, to have difficulty performing verbal working memory tasks. Clinical cases involving patients diagnosed with aphasia and specific language impairments have highlighted challenges related to retaining and manipulating auditory information. For example, those who sustain damage specifically within their left inferior frontal gyrus often struggle with tasks involving phonological rehearsal and verbal working memory activities, and therefore, they tend to perform poorly in tasks that require manipulation or repetition of verbal stimuli ( Saffran, 1997 ; Caplan and Waters, 2005 ).
The visuospatial sketchpad is engaged in the temporary retention and manipulation of visuospatial facts, including mental pictures, spatial associations, and object placements ( Miyake et al., 2001 ). The visuospatial sketchpad is localized to the right hemisphere, including the occipital lobe, parietal and frontal areas ( Osaka et al., 2007 ). Ren et al. (2019) identified the localization of the visuospatial sketchpad, and these areas were the right infero-lateral prefrontal cortex, lateral pre-motor cortices, right inferior parietal cortex, and the dorsolateral occipital cortices ( Burbaud et al., 1999 ; Salvato et al., 2021 ). Moreover, the posterior parietal cortex and the intraparietal sulcus have been implicated in spatial working memory ( Xu and Chun, 2006 ). Additionally, some evidence is available for an increase in brain regions associated with the visuospatial sketchpad during tasks involving mental imagery and spatial processing. Neuroimaging studies have revealed increased neural activation in some regions of the parietal cortex, mainly the superior and posterior parietal cortex, while performing mental rotation tasks ( Cohen et al., 1996 ; Kosslyn et al., 1997 ). However, further research is needed to better understand the visuospatial working memory and its integration with other cognitive processes ( Baddeley, 2003 ). Lesions to the regions involving the visuospatial sketchpad can have detrimental effects on visuospatial working memory tasks. Individuals with lesions to the posterior parietal cortex may exhibit deficits in mental rotation tasks and may be unable to mentally manipulate the visuospatial representation ( Buiatti et al., 2011 ). Moreover, studies concerning lesions have shown that damage to the parietal cortex can result in short-term deficits in visuospatial memory ( Shafritz et al., 2002 ). Damage to the occipital cortex can lead to performance impairments in tasks that require the generation and manipulation of mental visual images ( Moro et al., 2008 ).
The fourth component of the working memory, termed episodic buffer, was proposed by Baddeley (2000) . The episodic buffer is a multidimensional but essentially passive store that can hold a limited number of chunks, store bound features, and make them available to conscious awareness ( Baddeley et al., 2010 ; Hitch et al., 2019 ). Although research has suggested that episodic buffer is localized to the hippocampus ( Berlingeri et al., 2008 ) or the inferior lateral parietal cortex, it is thought to be not dependent on a single anatomical structure but instead can be influenced by the subsystems of working memory, long term memory, and even through perception ( Vilberg and Rugg, 2008 ; Baddeley et al., 2010 ). The episodic buffer provides a crucial link between the attentional central executive and the multidimensional information necessary for the operation of working memory ( Baddeley et al., 2011 ; Gelastopoulos et al., 2019 ).
The interdependence of the working memory modules, namely the phonological loop and visuospatial sketchpad, co-relates with other cognitive processes, for instance, spatial cognition and attention allocation ( Repovs and Baddeley, 2006 ). It has been found that the prefrontal cortex (PFC) and posterior parietal cortex (PPC) have a crucial role in several aspects of spatial cognition, such as the maintenance of spatially oriented attention and motor intentions ( Jerde and Curtis, 2013 ). The study by Sellers et al. (2016) and the review by Ikkai and Curtis (2011) posits that other brain areas could use the activity in PFC and PPC as a guide and manifest outputs to guide attention allocation, spatial memory, and motor planning. Moreover, research indicates that verbal information elicits an activation response in the left ventrolateral prefrontal cortex (VLPFC) when retained in the phonological loop, while visuospatial information is represented by a corresponding level of activity within the right homolog region ( Narayanan et al., 2005 ; Wolf et al., 2006 ; Emch et al., 2019 ). Specifically, the study by Yang et al. (2022) investigated the roles of two regions in the brain, the right inferior frontal gyrus (rIFG) and the right supra-marginal gyrus (rSMG), as they relate to spatial congruency in visual working memory tasks. A change detection task with online repetitive transcranial magnetic stimulation applied concurrently at both locations during high visual WM load conditions determined that rIFG is involved in actively repositioning the location of objects. At the same time, rSMG is engaged in passive perception of the stability of the location of objects.
Recent academic studies have found evidence to support the development of a new working memory model known as the state-based model ( D’Esposito and Postle, 2015 ). This theoretical model proposes that the allocation of attention toward internal representations permits short-term retention within working memory ( Ghaleh et al., 2019 ). The state-based model consists of two main categories: activated LTM models and sensorimotor recruitment models; the former largely focuses upon symbolic stimuli categorized under semantic aspects, while the latter has typically been applied to more perceptual tasks in experiments. This framework posits that prioritization through regulating cognitive processes provides insight into various characteristics across different activity types, including capacity limitations, proactive interference, etcetera ( D’Esposito and Postle, 2015 ). For example, the paper by Ghaleh et al. (2019) provides evidence for two separate mechanisms involved in maintenance of auditory information in verbal working memory: an articulatory rehearsal mechanism that relies more heavily on left sensorimotor areas and a non-articulatory maintenance mechanism that critically relies on left superior temporal gyrus (STG). These findings support the state-based model’s proposal that attentional allocation is necessary for short-term retention in working memory.
State-based models were found to be consistent with the suggested storage mechanism as they do not require representation transfer from one dedicated buffer type; research has demonstrated that any population of neurons and synapses may serve as such buffers ( Maass and Markram, 2002 ; Postle, 2006 ; Avraham et al., 2017 ). The review by D’Esposito and Postle (2015) examined the evidence to determine whether a persistent neural activity, synaptic mechanisms, or a combination thereof support representations maintained during working memory. Numerous neural mechanisms have been hypothesized to support the short-term retention of information in working memory and likely operate in parallel ( Sreenivasan et al., 2014 ; Kamiński and Rutishauser, 2019 ).
Persistent neural activity is the neural mechanism by which information is temporarily maintained ( Ikkai and Curtis, 2011 ; Panzeri et al., 2023 ). Recent review by Curtis and Sprague (2021) has focused on the notion that persistent neural activity is a fundamental mechanism for memory storage and have provided two main arcs of explanation. The first arc, mainly underpinned by empirical evidence from prefrontal cortex (PFC) neurophysiology experiments and computational models, posits that PFC neurons exhibit sustained firing during working memory tasks, enabling them to store representations in their active state ( Thuault et al., 2013 ). Intrinsic persistent firing in layer V neurons in the medial PFC has been shown to be regulated by HCN1 channels, which contribute to the executive function of the PFC during working memory episodes ( Thuault et al., 2013 ). Additionally, research has also found that persistent neural firing could possibly interact with theta periodic activity to sustain each other in the medial temporal, prefrontal, and parietal regions ( Düzel et al., 2010 ; Boran et al., 2019 ). The second arc involves advanced neuroimaging approaches which have, more recently, enabled researchers to decode content stored within working memories across distributed regions of the brain, including parts of the early visual cortex–thus extending this framework beyond just isolated cortical areas such as the PFC. There is evidence that suggests simple, stable, persistent activity among neurons in stimulus-selective populations may be a crucial mechanism for sustaining WM representations ( Mackey et al., 2016 ; Kamiński et al., 2017 ; Curtis and Sprague, 2021 ).
Badre (2008) discussed the functional organization of the PFC. The paper hypothesized that the rostro-caudal gradient of a function in PFC supported a control hierarchy, whereas posterior to anterior PFC mediated progressively abstract, higher-order controls ( Badre, 2008 ). However, this outlook proposed by Badre (2008) became outdated; the paper by Badre and Nee (2018) presented an updated look at the literature on hierarchical control. This paper supports neither a unitary model of lateral frontal function nor a unidimensional abstraction gradient. Instead, separate frontal networks interact via local and global hierarchical structures to support diverse task demands. This updated perspective is supported by recent studies on the hierarchical organization of representations within the lateral prefrontal cortex (LPFC) and the progressively rostral areas of the LPFC that process/represent increasingly abstract information, facilitating efficient and flexible cognition ( Thomas Yeo et al., 2011 ; Nee and D’Esposito, 2016 ). This structure allows the brain to access increasingly abstract action representations as required ( Nee and D’Esposito, 2016 ). It is supported by fMRI studies showing an anterior-to-posterior activation movement when tasks become more complex. Anatomical connectivity between areas also supports this theory, such as Area 10, which has projections back down to Area 6 but not vice versa.
Finally, studies confirm that different regions serve different roles along a hierarchy leading toward goal-directed behavior ( Badre and Nee, 2018 ). The paper by Postle (2015) exhibits evidence of activity in the prefrontal cortex that reflects the maintenance of high-level representations, which act as top-down signals, and steer the circulation of neural pathways across brain networks. The PFC is a source of top-down signals that influence processing in the posterior and subcortical regions ( Braver et al., 2008 ; Friedman and Robbins, 2022 ). These signals either enhance task-relevant information or suppress irrelevant stimuli, allowing for efficient yet effective search ( D’Esposito, 2007 ; D’Esposito and Postle, 2015 ; Kerzel and Burra, 2020 ). The study by Ratcliffe et al. (2022) provides evidence of the dynamic interplay between executive control mechanisms in the frontal cortex and stimulus representations held in posterior regions for working memory tasks. Moreover, the review by Herry and Johansen (2014) discusses the neural mechanisms behind actively maintaining task-relevant information in order for a person to carry out tasks and goals effectively. This review of data and research suggests that working memory is a multi-component system allowing for both the storage and processing of temporarily active representations. Neural activity throughout the brain can be differentially enhanced or suppressed based on context through top-down signals emanating from integrative areas such as PFC, parietal cortex, or hippocampus to actively maintain task-relevant information when it is not present in the environment ( Herry and Johansen, 2014 ; Kerzel and Burra, 2020 ).
In addition, Yu et al. (2022) examined how brain regions from the ventral stream pathway to the prefrontal cortex were activated during working memory (WM) gate opening and closing. They defined gate opening as the switch from maintenance to updating and gate closing as the switch from updating to maintenance. The data suggested that cognitive branching increases during the WM gating process, thus correlating the gating process and an information approach to the PFC function. The temporal cortices, lingual gyrus (BA19), superior frontal gyri including frontopolar cortices, and middle and inferior parietal regions are involved in processes of estimating whether a response option available will be helpful for each case. During gate closing, on the other hand, medial and superior frontal regions, which have been associated with conflict monitoring, come into play, as well as orbitofrontal and dorsolateral prefrontal processing at later times when decreasing activity resembling stopping or downregulating cognitive branching has occurred, confirming earlier theories about these areas being essential for estimation of usefulness already stored within long-term memories ( Yu et al., 2022 ).
Declarative and non-declarative memory
The distinctions between declarative and non-declarative memory are often based on the anatomical features of medial temporal lobe regions, specifically those involving the hippocampus ( Squire and Zola, 1996 ; Squire and Wixted, 2011 ). In the investigation of systems implicated in the process of learning and memory formation, it has been posited that the participation of the hippocampus is essential for the acquisition of declarative memories ( Eichenbaum and Cohen, 2014 ). In contrast, a comparatively reduced level of hippocampal involvement may suffice for non-declarative memories ( Squire and Zola, 1996 ; Williams, 2020 ).
Declarative memory (explicit) pertains to knowledge about facts and events. This type of information can be consciously retrieved with effort or spontaneously recollected without conscious intention ( Dew and Cabeza, 2011 ). There are two types of declarative memory: Episodic and Semantic. Episodic memory is associated with the recollection of personal experiences. It involves detailed information about events that happened in one’s life. Semantic memory refers to knowledge stored in the brain as facts, concepts, ideas, and objects; this includes language-related information like meanings of words and mathematical symbol values along with general world knowledge (e.g., capitals of countries) ( Binder and Desai, 2011 ). The difference between episodic and semantic memory is that when one retrieves episodic memory, the experience is known as “remembering”; when one retrieves information from semantic memory, the experience is known as “knowing” ( Tulving, 1985 ; Dew and Cabeza, 2011 ). The hippocampus, medial temporal lobe, and the areas in the diencephalon are implicated in declarative memory ( Richter-Levin and Akirav, 2003 ; Derner et al., 2020 ). The ventral parietal cortex (VPC) is involved in declarative memory processes, specifically episodic memory retrieval ( Henson et al., 1999 ; Davis et al., 2018 ). The evidence suggests that VPC and hippocampus is involved in the retrieval of contextual details, such as the location and timing of the event, and the information is critical for the formation of episodic memory ( Daselaar, 2009 ; Hutchinson et al., 2009 ; Wiltgen et al., 2010 ). The prefrontal cortex (PFC) is involved in the encoding (medial PFC) and retrieval (lateral PFC) of declarative memories, specifically in the integration of information across different sensory modalities ( Blumenfeld and Ranganath, 2007 ; Li et al., 2010 ). Research also suggests that the amygdala may modulate other brain regions involved with memory processing, thus, contributing to an enhanced recall of negative or positive experiences ( Hamann, 2001 ; Ritchey et al., 2008 ; Sendi et al., 2020 ). Maintenance of the integrity of hippocampal circuitry is essential for ensuring that episodic memory, along with spatial and temporal context information, can be retained in short-term or long-term working memory beyond 15 min ( Ito et al., 2003 ; Rasch and Born, 2013 ). Moreover, studies have suggested that the amygdala plays a vital role in encoding and retrieving explicit memories, particularly those related to emotionally charged stimuli which are supported by evidence of correlations between hippocampal activity and amygdala modulation during memory formation ( Richter-Levin and Akirav, 2003 ; Qasim et al., 2023 ).
Current findings in neuroimaging studies assert that a vast array of interconnected brain regions support semantic memory ( Binder and Desai, 2011 ). This network merges information sourced from multiple senses alongside different cognitive faculties necessary for generating abstract supramodal views on various topics stored within our consciousness. Modality-specific sensory, motor, and emotional system within these brain regions serve specialized tasks like language comprehension, while larger areas of the brain, such as the inferior parietal lobe and most of the temporal lobe, participate in more generalized interpretation tasks ( Binder and Desai, 2011 ; Kuhnke et al., 2020 ). These regions lie at convergences of multiple perceptual processing streams, enabling increasingly abstract, supramodal representations of perceptual experience that support a variety of conceptual functions, including object recognition, social cognition, language, and the remarkable human capacity to remember the past and imagine the future ( Binder and Desai, 2011 ; Binney et al., 2016 ). The following section will discuss the processes underlying memory consolidation and storage within declarative memory.
Non-declarative (implicit) memories refer to unconscious learning through experience, such as habits and skills formed from practice rather than memorizing facts; these are typically acquired slowly and automatically in response to sensory input associated with reward structures or prior exposure within our daily lives ( Kesner, 2017 ). Non-declarative memory is a collection of different phenomena with different neural substrates rather than a single coherent system ( Camina and Güell, 2017 ). It operates by similar principles, depending on local changes to a circumscribed brain region, and the representation of these changes is unavailable to awareness ( Reber, 2008 ). Non-declarative memory encompasses a heterogenous collection of abilities, such as associative learning, skills, and habits (procedural memory), priming, and non-associative learning ( Squire and Zola, 1996 ; Camina and Güell, 2017 ). Studies have concluded that procedural memory for motor skills depends upon activity in diverse set areas such as the motor cortex, striatum, limbic system, and cerebellum; similarly, perceptual skill learning is thought to be associated with sensory cortical activation ( Karni et al., 1998 ; Mayes, 2002 ). Research suggests that mutual connections between brain regions that are active together recruit special cells called associative memory cells ( Wang et al., 2016 ; Wang and Cui, 2018 ). These cells help integrate, store, and remember related information. When activated, these cells trigger the recall of memories, leading to behaviors and emotional responses. This suggests that co-activated brain regions with these mutual connections are where associative memories are formed ( Wang et al., 2016 ; Wang and Cui, 2018 ). Additionally, observational data reveals that priming mechanisms within distinct networks, such as the “repetition suppression” effect observed in visual cortical areas associated with sensory processing and in the prefrontal cortex for semantic priming, are believed to be responsible for certain forms of conditioning and implicit knowledge transfer experiences exhibited by individuals throughout their daily lives ( Reber, 2008 ; Wig et al., 2009 ; Camina and Güell, 2017 ). However, further research is needed to better understand the mechanisms of consolidation in non-declarative memory ( Camina and Güell, 2017 ).
The process of transforming memory into stable, long-lasting from a temporary, labile memory is known as memory consolidation ( McGaugh, 2000 ). Memory formation is based on the change in synaptic connections of neurons representing the memory. Encoding causes synaptic Long-Term potentiation (LTP) or Long-Term depression (LTD) and induces two consolidation processes. The first is synaptic or cellular consolidation which involves remodeling synapses to produce enduring changes. Cellular consolidation is a short-term process that involves stabilizing the neural trace shortly after learning via structural brain changes in the hippocampus ( Lynch, 2004 ). The second is system consolidation, which builds on synaptic consolidation where reverberating activity leads to redistribution for long-term storage ( Mednick et al., 2011 ; Squire et al., 2015 ). System consolidation is a long-term process during which memories are gradually transferred to and integrated with cortical neurons, thus promoting their stability over time. In this way, memories are rendered less susceptible to forgetting. Hebb postulated that when two neurons are repeatedly activated simultaneously, they become more likely to exhibit a coordinated firing pattern of activity in the future ( Langille, 2019 ). This proposed enduring change in synchronized neuronal activation was consequently termed cellular consolidation ( Bermudez-Rattoni, 2010 ).
The following sections of this paper incorporate a more comprehensive investigation into various essential procedures connected with memory consolidation- namely: long-term potentiation (LTP), long-term depression (LTD), system consolidation, and cellular consolidation. Although these mechanisms have been presented briefly before this paragraph, the paper aims to offer greater insight into each process’s function within the individual capacity and their collective contribution toward memory consolidation.
Synaptic plasticity mechanisms implicated in memory stabilization
Long-Term Potentiation (LTP) and Long-Term Depression (LTP) are mechanisms that have been implicated in memory stabilization. LTP is an increase in synaptic strength, whereas LTD is a decrease in synaptic strength ( Ivanco, 2015 ; Abraham et al., 2019 ).
Long-Term Potentiation (LTP) is a phenomenon wherein synaptic strength increases persistently due to brief exposures to high-frequency stimulation ( Lynch, 2004 ). Studies of Long-Term Potentiation (LTP) have led to an understanding of the mechanisms behind synaptic strengthening phenomena and have provided a basis for explaining how and why strong connections between neurons form over time in response to stimuli.
The NMDA receptor-dependent LTP is the most commonly described LTP ( Bliss and Collingridge, 1993 ; Luscher and Malenka, 2012 ). In this type of LTP, when there is high-frequency stimulation, the presynaptic neuron releases glutamate, an excitatory neurotransmitter. Glutamate binds to the AMPA receptor on the postsynaptic neuron, which causes the neuron to fire while opening the NMDA receptor channel. The opening of an NMDA channel elicits a calcium ion influx into the postsynaptic neuron, thus initiating a series of phosphorylation events as part of the ensuing molecular cascade. Autonomously phosphorylated CaMKII and PKC, both actively functional through such a process, have been demonstrated to increase the conductance of pre-existing AMPA receptors in synaptic networks. Additionally, this has been shown to stimulate the introduction of additional AMPA receptors into synapses ( Malenka and Nicoll, 1999 ; Lynch, 2004 ; Luscher and Malenka, 2012 ; Bailey et al., 2015 ).
There are two phases of LTP: the early phase and the late phase. It has been established that the early phase LTP (E-LTP) does not require RNA or protein synthesis; therefore, its synaptic strength will dissipate in minutes if late LTP does not stabilize it. On the contrary, late-phase LTP (L-LTP) can sustain itself over a more extended period, from several hours to multiple days, with gene transcription and protein synthesis in the postsynaptic cell ( Frey and Morris, 1998 ; Orsini and Maren, 2012 ). The strength of presynaptic tetanic stimulation has been demonstrated to be a necessary condition for the activation of processes leading to late LTP ( Luscher and Malenka, 2012 ; Bailey et al., 2015 ). This finding is supported by research examining synaptic plasticity, notably Eric Kandel’s discovery that CREB–a transcription factor–among other cytoplasmic and nuclear molecules, are vital components in mediating molecular changes culminating in protein synthesis during this process ( Kaleem et al., 2011 ; Kandel et al., 2014 ). Further studies have shown how these shifts ultimately lead to AMPA receptor stabilization at post-synapses facilitating long-term potentiation within neurons ( Luscher and Malenka, 2012 ; Bailey et al., 2015 ).
The “synaptic tagging and capture hypothesis” explains how a weak event of tetanization at synapse A can transform to late-LTP if followed shortly by the strong tetanization of a different, nearby synapse on the same neuron ( Frey and Morris, 1998 ; Redondo and Morris, 2011 ; Okuda et al., 2020 ; Park et al., 2021 ). During this process, critical plasticity-related proteins (PRPs) are synthesized, which stabilize their own “tag” and that from the weaker synaptic activity ( Moncada et al., 2015 ). Recent evidence suggests that calcium-permeable AMPA receptors (CP-AMPARs) are involved in this form of heterosynaptic metaplasticity ( Park et al., 2018 ). The authors propose that the synaptic activation of CP-AMPARs triggers the synthesis of PRPs, which are then engaged by the weak induction protocol to facilitate LTP on the independent input. The paper also suggests that CP-AMPARs are required during the induction of LTP by the weak input for the full heterosynaptic metaplastic effect to be observed ( Park et al., 2021 ). Additionally, it has been further established that catecholamines such as dopamine plays an integral part in memory persistence by inducing PRP synthesis ( Redondo and Morris, 2011 ; Vishnoi et al., 2018 ). Studies have found that dopamine release in the hippocampus can enhance LTP and improve memory consolidation ( Lisman and Grace, 2005 ; Speranza et al., 2021 ).
Investigations into neuronal plasticity have indicated that synaptic strength alterations associated with certain forms of learning and memory may be analogous to those underlying Long-Term Potentiation (LTP). Research has corroborated this notion, demonstrating a correlation between these two phenomena ( Lynch, 2004 ). The three essential properties of Long-Term Potentiation (LTP) that have been identified are associativity, synapse specificity, and cooperativity ( Kandel and Mack, 2013 ). These characteristics provide empirical evidence for the potential role of LTP in memory formation processes. Specifically, associativity denotes the amplification of connections when weak stimulus input is paired with a powerful one; synapse specificity posits that this potentiating effect only manifests on synaptic locations exhibiting coincidental activity within postsynaptic neurons, while cooperativity suggests stimulated neuron needs to attain an adequate threshold of depolarization before LTP can be induced again ( Orsini and Maren, 2012 ).
There is support for the idea that memories are encoded by modification of synaptic strengths through cellular mechanisms such as LTP and LTD ( Nabavi et al., 2014 ). The paper by Nabavi et al. (2014) shows that fear conditioning, a type of associative memory, can be inactivated and reactivated by LTD and LTP, respectively. The findings of the paper support a causal link between these synaptic processes and memory. Moreover, the paper suggests that LTP is used to form neuronal assemblies that represent a memory, and LTD could be used to disassemble them and thereby inactivate a memory ( Nabavi et al., 2014 ). Hippocampal LTD has been found to play an essential function in regulating synaptic strength and forming memories, such as long-term spatial memory ( Ge et al., 2010 ). However, it is vital to bear in mind that studies carried out on LTP exceed those done on LTD; hence the literature on it needs to be more extensive ( Malenka and Bear, 2004 ; Nabavi et al., 2014 ).
Cellular consolidation and memory
For an event to be remembered, it must form physical connections between neurons in the brain, which creates a “memory trace.” This memory trace can then be stored as long-term memory ( Langille and Brown, 2018 ). The formation of a memory engram is an intricate process requiring neuronal depolarization and the influx of intracellular calcium ( Mank and Griesbeck, 2008 ; Josselyn et al., 2015 ; Xu et al., 2017 ). This initiation leads to a cascade involving protein transcription, structural and functional changes in neural networks, and stabilization during the quiescence period, followed by complete consolidation for its success. Interference from new learning events or disruption caused due to inhibition can abort this cycle leading to incomplete consolidation ( Josselyn et al., 2015 ).
Cyclic-AMP response element binding protein (CREB) has been identified as an essential transcription factor for memory formation ( Orsini and Maren, 2012 ). It regulates the expression of PRPs and enhances neuronal excitability and plasticity, resulting in changes to the structure of cells, including the growth of dendritic spines and new synaptic connections. Blockage or enhancement of CREB in certain areas can affect subsequent consolidation at a systems level–decreasing it prevents this from occurring, while aiding its presence allows even weak learning conditions to produce successful memory formation ( Orsini and Maren, 2012 ; Kandel et al., 2014 ).
Strengthening weakly encoded memories through the synaptic tagging and capture hypothesis may play an essential role in cellular consolidation. Retroactive memory enhancement has also been demonstrated in human studies, mainly when items are initially encoded with low strength but later paired with shock after consolidation ( Dunsmoor et al., 2015 ). The synaptic tagging and capture theory (STC) and its extension, the behavioral tagging hypothesis (BT), have both been used to explain synaptic specificity and the persistence of plasticity ( Moncada et al., 2015 ). STC proposed that electrophysiological activity can induce long-term changes in synapses, while BT postulates similar effects of behaviorally relevant neuronal events on learning and memory models. This hypothesis proposes that memory consolidation relies on combining two distinct processes: setting a “learning tag” and synthesizing plasticity-related proteins ( De novo protein synthesis, increased CREB levels, and substantial inputs to nearby synapses) at those tagged sites. BT explains how it is possible for event episodes with low-strength inputs or engagements can be converted into lasting memories ( Lynch, 2004 ; Moncada et al., 2015 ). Similarly, the emotional tagging hypothesis posits that the activation of the amygdala in emotionally arousing events helps to mark experiences as necessary, thus enhancing synaptic plasticity and facilitating transformation from transient into more permanent forms for encoding long-term memories ( Richter-Levin and Akirav, 2003 ; Zhu et al., 2022 ).
Cellular consolidation, the protein synthesis-dependent processes observed in rodents that may underlie memory formation and stabilization, has been challenging to characterize in humans due to the limited ability to study it directly ( Bermudez-Rattoni, 2010 ). Additionally, multi-trial learning protocols commonly used within human tests as opposed to single-trial experiments conducted with non-human subjects suggest there could be interference from subsequent information that impedes individual memories from being consolidated reliably. This raises important questions regarding how individuals can still form strong and long-lasting memories when exposed to frequent stimuli outside controlled laboratory conditions. Although this phenomenon remains undiscovered by science, it is of utmost significance for gaining a deeper understanding of our neural capacities ( Genzel and Wixted, 2017 ).
The establishment of distributed memory traces requires a narrow temporal window following the initial encoding process, during which cellular consolidation occurs ( Nader and Hardt, 2009 ). Once this period ends and consolidation has been completed, further protein synthesis inhibition or pharmacological disruption will be less effective at altering pre-existing memories and interfering with new learning due to the stabilization of the trace in its new neuronal network connections ( Nader and Hardt, 2009 ). Thus, systems consolidation appears critical for the long-term maintenance of memory within broader brain networks over extended periods after their formation ( Bermudez-Rattoni, 2010 ).
System consolidation and memory
Information is initially stored in both the hippocampus and neocortex ( Dudai et al., 2015 ). The hippocampus subsequently guides a gradual process of reorganization and stabilization whereby information present within the neocortex becomes autonomous from that in the hippocampal store. Scholars have termed this phenomenon “standard memory consolidation model” or “system consolidation” ( Squire et al., 2015 ).
The Standard Model suggests that information acquired during learning is simultaneously stored in both the hippocampus and multiple cortical modules. Subsequently, it posits that over a period of time which may range from weeks to months or longer, the hippocampal formation directs an integration process by which these various elements become enclosed into single unified structures within the cortex ( Gilboa and Moscovitch, 2021 ; Howard et al., 2022 ). These newly learned memories are then assimilated into existing networks without interference or compression when necessary ( Frankland and Bontempi, 2005 ). It is important to note that memory engrams already exist within cortical networks during encoding. They only need strengthening through links enabled by hippocampal assistance-overtime allowing remote memory storage without reliance on the latter structure. Data appears consistent across studies indicating that both AMPA-and NMDA receptor-dependent “tagging” processes occurring within the cortex are essential components of progressive rewiring, thus enabling longer-term retention ( Takeuchi et al., 2014 ; Takehara-Nishiuchi, 2020 ).
Recent studies have additionally demonstrated that the rate of system consolidation depends on an individual’s ability to relate new information to existing networks made up of connected neurons, popularly known as “schemas” ( Robin and Moscovitch, 2017 ). In situations where prior knowledge is present and cortical modules are already connected at the outset of learning, it has been observed that a hippocampal-neocortical binding process occurs similarly to when forming new memories ( Schlichting and Preston, 2015 ). The proposed framework involves the medial temporal lobe (MTL), which is involved in acquiring new information and binds different aspects of an experience into a single memory trace. In contrast, the medial prefrontal cortex (mPFC) integrates this information with the existing knowledge ( Zeithamova and Preston, 2010 ; van Kesteren et al., 2012 ). During consolidation and retrieval, MTL is involved in replaying memories to the neocortex, where they are gradually integrated with existing knowledge and schemas and help retrieve memory traces. During retrieval, the mPFC is thought to use existing knowledge and schemas to guide retrieval and interpretation of memory. This may involve the assimilation of newly acquired information into existing cognitive schemata as opposed to the comparatively slow progression of creating intercortical connections ( Zeithamova and Preston, 2010 ; van Kesteren et al., 2012 , 2016 ).
Medial temporal lobe structures are essential for acquiring new information and necessary for autobiographical (episodic) memory ( Brown et al., 2018 ). The consolidation of autobiographical memories depends on a distributed network of cortical regions. Brain areas such as entorhinal, perirhinal, and parahippocampal cortices are essential for learning new information; however, they have little impact on the recollection of the past ( Squire et al., 2015 ). The hippocampus is a region of the brain that forms episodic memories by linking multiple events to create meaningful experiences ( Cooper and Ritchey, 2019 ). It receives information from all areas of the association cortex and cingulate cortex, subcortical regions via the fornix, as well as signals originating within its entorhinal cortex (EC) and amygdala regarding emotionally laden or potentially hazardous stimuli ( Sorensen, 2009 ). Such widespread connectivity facilitates the construction of an accurate narrative underpinning each remembered episode, transforming short-term into long-term recollections ( Richter-Levin and Akirav, 2003 ).
Researchers have yet to establish a consensus regarding where semantic memory information is localized within the brain ( Roldan-Valadez et al., 2012 ). Some proponents contend that such knowledge is lodged within perceptual and motor systems, triggered when we initially associate with a given object. This point of view is supported by studies highlighting how neural activity occurs initially in the occipital cortex, followed by left temporal lobe involvement during processing and pertinent contributions to word selection/retrieval via activation of left inferior frontal cortices ( Patterson et al., 2007 ). Moreover, research indicates elevated levels of fusiform gyrus engagement (a ventral surface region encompassing both temporal lobes) occurring concomitantly with verbal comprehension initiatives, including reading and naming tasks ( Patterson et al., 2007 ).
Research suggests that the hippocampus is needed for a few years after learning to support semantic memory (factual information), yet, it is not needed for the long term ( Squire et al., 2015 ). However, some forms of memory remain dependent on the hippocampus, such as the retrieval of spatial memory ( Wiltgen et al., 2010 ). Similarly, the Multiple-trace theory ( Moscovitch et al., 2006 ), also known as the transformation hypothesis ( Winocur and Moscovitch, 2011 ), posits that hippocampal engagement is necessary for memories that retain contextual detail such as episodic memories. Consolidation of memories into the neocortex is theorized to involve a loss of specific finer details, such as temporal and spatial information, in addition to contextual elements. This transition ultimately results in an evolution from episodic memory toward semantic memory, which consists mainly of gist-based facts ( Moscovitch et al., 2006 ).
Sleep and memory consolidation
Sleep is an essential physiological process crucial to memory consolidation ( Siegel, 2001 ). Sleep is divided into two stages: Non-rapid Eye Movement (NREM) sleep and Rapid Eye Movement (REM) sleep. NREM sleep is divided into three stages: N1, N2, and N3 (AKA Slow Wave Sleep or SWS) ( Rasch and Born, 2013 ). Each stage displays unique oscillatory patterns and phenomena responsible for consolidating memories in distinct ways. The first stage, or N1 sleep, is when an individual transitions between wakefulness and sleep. This type of sleep is characterized by low-amplitude, mixed-frequency brain activity. N1 sleep is responsible for the initial encoding of memories ( Rasch and Born, 2013 ). The second stage, or N2 sleep, is characterized by the occurrence of distinct sleep spindles and K-complexes in EEG. N2 is responsible for the consolidation of declarative memories ( Marshall and Born, 2007 ). The third stage of sleep N3, also known as slow wave sleep (SWS), is characterized by low-frequency brain activity, slow oscillations, and high amplitude. The slow oscillations which define the deepest stage of sleep are trademark rhythms of NREM sleep. These slow oscillations are delta waves combined to indicate slow wave activity (SWA), which is implicated in memory consolidation ( Tononi and Cirelli, 2003 ; Stickgold, 2005 ; Kim et al., 2019 ). Sleep spindles are another trademark defining NREM sleep ( Stickgold, 2005 ). Ripples are high-frequency bursts, and when combined with irregularly occurring sharp waves (high amplitude), they form the sharp-wave ripple (SWR). These spindles and the SWRs coordinate the reactivation and redistribution of hippocampus-dependent memories to neocortical sites ( Ngo et al., 2020 ; Girardeau and Lopes-dos-Santos, 2021 ). The third stage is also responsible for the consolidation of procedural memories, such as habits and motor skills ( Diekelmann and Born, 2010 ). During SWS, there is minimal cholinergic activity and intermediate noradrenergic activity ( Datta and MacLean, 2007 ).
Finally, the fourth stage of sleep is REM sleep, characterized by phasic REMs and muscle atonia ( Reyes-Resina et al., 2021 ). During REM sleep, there is high cholinergic activity, serotonergic and noradrenergic activity are at a minimum, and high theta activity ( Datta and MacLean, 2007 ). REM sleep is also characterized by local increases in plasticity-related immediate-early gene activity, which might favor the subsequent synaptic consolidation of memories in the cortex ( Ribeiro, 2007 ; Diekelmann and Born, 2010 ; Reyes-Resina et al., 2021 ). The fourth stage of sleep is responsible for the consolidation of emotional memories and the integration of newly acquired memories into existing knowledge structures ( Rasch and Born, 2013 ). Studies indicate that the cholinergic system plays an imperative role in modifying these processes by toggling the entire thalamo-cortico-hippocampal network between distinct modes, namely high Ach encoding mode during active wakefulness and REM sleep and low Ach consolidation mode during quiet wakefulness and NREM sleep ( Bergmann and Staresina, 2017 ; Li et al., 2020 ). Consequently, improving neocortical hippocampal communication results in efficient memory encoding/synaptic plasticity, whereas hippocampo-neocortical interactions favor better systemic memory consolidation ( Diekelmann and Born, 2010 ).
The dual process hypothesis of memory consolidation posits that SWS facilitates declarative, hippocampus-dependent memory, whereas REM sleep facilitates non-declarative hippocampus-independent memory ( Maquet, 2001 ; Diekelmann and Born, 2010 ). On the other hand, the sequential hypothesis states that different sleep stages play a sequential role in memory consolidation. Memories are encoded during wakefulness, consolidated during NREM sleep, and further processed and integrated during REM sleep ( Rasch and Born, 2013 ). However, there is evidence present that contradicts the sequential hypothesis. A study by Goerke et al. (2013) found that declarative memories can be consolidated during REM sleep, suggesting that the relationship between sleep stages and memory consolidation is much more complex than a sequential model. Moreover, other studies indicate the importance of coordinating specific sleep phases with learning moments for optimal memory retention. This indicates that the timing of sleep has more influence than the specific sleep stages ( Gais et al., 2006 ). The active system consolidation theory suggests that an active consolidation process results from the selective reactivation of memories during sleep; the brain selectively reactivates newly encoded memories during sleep, which enhances and integrates them into the network of pre-existing long-term memories ( Born et al., 2006 ; Howard et al., 2022 ). Research has suggested that slow-wave sleep (SWS) and rapid eye movement (REM) sleep have complementary roles in memory consolidation. Declarative and non-declarative memories benefiting differently depending on which sleep stage they rely on ( Bergmann and Staresina, 2017 ). Specifically, during SWS, the brain actively reactivates and reorganizes hippocampo-neocortical memory traces as part of system consolidation. Following this, REM sleep is crucial for stabilizing these reactivated memory traces through synaptic consolidation. While SWS may initiate early plastic processes in hippocampo-neocortical memory traces by “tagging” relevant neocortico-neocortical synapses for later consolidation ( Frey and Morris, 1998 ), long-term plasticity requires subsequent REM sleep ( Rasch and Born, 2007 , 2013 ).
The active system consolidation hypothesis is not the only mechanism proposed for memory consolidation during sleep. The synaptic homeostasis hypothesis proposes that sleep is necessary for restoring synaptic homeostasis, which is challenged by synaptic strengthening triggered by learning during wake and synaptogenesis during development ( Tononi and Cirelli, 2014 ). The synaptic homeostasis hypothesis assumes consolidation is a by-product of the global synaptic downscaling during sleep ( Puentes-Mestril and Aton, 2017 ). The two models are not mutually exclusive, and the hypothesized processes probably act in concert to optimize the memory function of sleep ( Diekelmann and Born, 2010 ).
Non-rapid eye movement sleep plays an essential role in the systems consolidation of memories, with evidence showing that different oscillations are involved in this process ( Düzel et al., 2010 ). With an oscillatory sequence initiated by a slow frontal cortex oscillation (0.5–1 Hz) traveling to the medial temporal lobe and followed by a sharp-wave ripple (SWR) in the hippocampus (100–200 Hz). Replay activity of memories can be measured during this oscillatory sequence across various regions, including the motor cortex and visual cortex ( Ji and Wilson, 2006 ; Eichenlaub et al., 2020 ). Replay activity of memory refers to the phenomenon where the hippocampus replays previously experienced events during sharp wave ripples (SWRs) and theta oscillations ( Zielinski et al., 2018 ). During SWRs, short, transient bursts of high-frequency oscillations occur in the hippocampus. During theta oscillations, hippocampal spikes are ordered according to the locations of their place fields during behavior. These sequential activities are thought to play a role in memory consolidation and retrieval ( Zielinski et al., 2018 ). The paper by Zielinski et al. (2018) suggests that coordinated hippocampal-prefrontal representations during replay and theta sequences play complementary and overlapping roles at different stages in learning, supporting memory encoding and retrieval, deliberative decision-making, planning, and guiding future actions.
Additionally, the high-frequency oscillations of SWR reactivate groups of neurons attributed to spatial information encoding to align synchronized activity across an array of neural structures, which results in distributed memory creation ( Swanson et al., 2020 ; Girardeau and Lopes-dos-Santos, 2021 ). Parallel to this process is slow oscillation or slow-wave activity within cortical regions, which reflects synced neural firing and allows regulation of synaptic weights, which is in accordance with the synaptic homeostasis hypothesis (SHY). The SHY posits that downscaling synaptic strengths help incorporate new memories by avoiding saturation of resources during extended periods–features validated by discoveries where prolonged wakefulness boosts amplitude while it diminishes during stretches of enhanced sleep ( Girardeau and Lopes-dos-Santos, 2021 ).
During REM sleep, the brain experiences “paradoxical” sleep due to the similarity in activity to wakefulness. This stage plays a significant role in memory processing. Theta oscillations which are dominant during REM sleep, are primarily observed in the hippocampus, and these are involved in memory consolidation ( Landmann et al., 2014 ). There has been evidence of coherence between theta oscillations in the hippocampus, medial frontal cortex, and amygdala, which support their involvement in memory consolidation ( Popa et al., 2010 ). During REM sleep, phasic events such as ponto-geniculo-occipital waves originating from the brainstem coordinate activity across various brain structures and may contribute to memory consolidation processes ( Rasch and Born, 2013 ). Research has suggested that sleep-associated consolidation may be mediated by the degree of overlap between new and already known material whereby, if the acquired information is similar to the information one has learned, it is more easily consolidated during sleep ( Tamminen et al., 2010 ; Sobczak, 2017 ).
In conclusion, understanding more about how the brains cycle through different stages of sleep, including specific wave patterns, offers valuable insight into the ability to store memories effectively. While NREM sleep is associated with SWRs and slow oscillations, facilitating memory consolidation and synaptic downscaling, REM sleep, characterized by theta oscillations and phasic events, contributes to memory reconsolidation and the coordination of activity across brain regions. By exploring the interactions between sleep stages, oscillations, and memory processes, one may learn more about how sleep impacts brain function and cognition in greater detail.
Century has passed since we addressed memory, and several notable findings have moved from bench-to-bedside research. Several cross-talks between multidiscipline have been encouraged. Nevertheless, further research is needed into neurobiological mechanisms of non-declarative memory, such as conditioning ( Gallistel and Balsam, 2014 ). Modern research indicates that structural change that encodes information is likely at the level of the synapse, and the computational mechanisms are implemented at the level of neural circuitry. However, it also suggests that intracellular mechanisms realized at the molecular level, such as micro RNAs, should not be discounted as potential mechanisms. However, further research is needed to study the molecular and structural changes brought on by implicit memory ( Gallistel and Balsam, 2014 ).
The contribution of non-human animal studies toward our understanding of memory processes cannot be understated; hence recognizing their value is vital for moving forward. While this paper predominantly focused on cognitive neuroscience perspectives, some articles cited within this paper were sourced from non-human animal studies providing fundamental groundwork and identification of critical mechanisms relevant to human memories. A need persists for further investigation—primarily with humans—which can validate existing findings from non-human animals. Moving forward, it is prudent for researchers to bridge the gap between animal and human investigations done while exploring parallels and exploring unique aspects of human memory processes. By integrating findings from both domains, one can gain a more comprehensive understanding of the complexities of memory and its underlying neural mechanisms. Such investigations will broaden the horizon of our memory process and answer the complex nature of memory storage.
This paper attempted to provide an overview and summarize memory and its processes. The paper focused on bringing the cognitive neuroscience perspective on memory and its processes. This may provide the readers with the understanding, limitations, and research perspectives of memory mechanisms.
Data availability statement
Author contributions.
SS and MKA: conceptualization, framework, and manuscript writing. AK: review and editing of the manuscript. All authors contributed to the article and approved the submitted version.
Acknowledgments
We gratefully thank students and Indian Institute of Technology Roorkee (IITR) office staff for their conditional and unconditional support. We also thank the Memory and Anxiety Research Group (MARG), IIT Roorkee for its constant support.
Funding Statement
MKA was supported by the F.I.G. grant (IITR/SRIC/2741). The funding agency had no role in the preparation of the manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
- Abraham W. C., Jones O. D., Glanzman D. L. (2019). Is plasticity of synapses the mechanism of long-term memory storage? NPJ Sci. Learn. 4 : 9 . 10.1038/s41539-019-0048-y [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Asthana M. K., Brunhuber B., Mühlberger A., Reif A., Schneider S., Herrmann M. J. (2015). Preventing the return of fear using reconsolidation update mechanisms depends on the met-allele of the brain derived neurotrophic factor val66met polymorphism. Int. J. Neuropsychopharmacol . 19 : yv137 . 10.1093/ijnp/pyv137 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Atkinson R. C., Shiffrin R. M. (1968). “ Human memory: a proposed system and its control processes ,” in Psychology of learning and motivation , Vol. 2 eds Spence K. W., Spence J. T. (Amsterdam: Elsevier; ), 89–195. 10.1016/S0079-7421(08)60422-3 [ CrossRef ] [ Google Scholar ]
- Avraham G., Leib R., Pressman A., Simo L. S., Karniel A., Shmuelof L., et al. (2017). State-based delay representation and its transfer from a game of pong to reaching and tracking. Eneuro 4 ENEURO.179–ENEURO.117. 10.1523/eneuro.0179-17.2017 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Awh E., Jonides J., Smith E. E., Schumacher E. H., Koeppe R. A., Katz S. (1996). Dissociation of storage and rehearsal in verbal working memory: evidence From Positron Emission Tomography. Psychol. Sci. 7 25–31. 10.1111/j.1467-9280.1996.tb00662.x [ CrossRef ] [ Google Scholar ]
- Baddeley A. (2000). The episodic buffer: a new component of working memory? Trends Cogn. Sci. 4 417–423. 10.1016/S1364-6613(00)01538-2 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Baddeley A. (2003). Working memory: looking back and looking forward. Nat. Rev. Neurosci. 4 829–839. 10.1038/nrn1201 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Baddeley A. D. (1986). Working memory. Oxford: Oxford University Press. [ Google Scholar ]
- Baddeley A. D., Allen R. J., Hitch G. J. (2011). Binding in visual working memory: the role of the episodic buffer. Neuropsychologia 49 1393–1400. 10.1016/j.neuropsychologia.2010.12.042 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Baddeley A. D., Hitch G. (1974). “ Working memory ,” in Psychology of learning and motivation , Vol. 8 ed. Bower G. A. (Amsterdam: Elsevier; ), 47–89. 10.1016/S0079-7421(08)60452-1 [ CrossRef ] [ Google Scholar ]
- Baddeley A. D., Logie R. H. (1999). “ Working memory: the multiple-component model ,” in Models of working memory , 1st Edn, eds Miyake A., Shah P. (Cambridge: Cambridge University Press; ), 28–61. 10.1017/CBO9781139174909.005 [ CrossRef ] [ Google Scholar ]
- Baddeley A., Allen R. J., Hitch G. J. (2010). Investigating the episodic buffer. Psychol. Belgica 50 223 . 10.5334/pb-50-3-4-223 [ CrossRef ] [ Google Scholar ]
- Badre D. (2008). Cognitive control, hierarchy, and the Rostro–caudal organization of the frontal lobes. Trends Cogn. Sci. 12 193–200. 10.1016/j.tics.2008.02.004 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Badre D., Nee D. E. (2018). Frontal cortex and the hierarchical control of behavior. Trends Cogn. Sci. 22 170–188. 10.1016/j.tics.2017.11.005 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Bailey C. H., Kandel E. R., Harris K. M. (2015). Structural components of synaptic plasticity and memory consolidation. Cold Spring Harb. Perspect. Biol. 7 : a021758 . 10.1101/cshperspect.a021758 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Bellfy L., Kwapis J. L. (2020). Molecular mechanisms of reconsolidation-dependent memory updating. Int. J. Mol. Sci. 21 : 6580 . 10.3390/ijms21186580 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Bergmann T. O., Staresina B. P. (2017). “ Neuronal oscillations and reactivation subserving memory consolidation ,” in Cognitive neuroscience of memory consolidation , eds Axmacher N., Rasch B. (Cham: Springer; ), 185–207. 10.1007/978-3-319-45066-7_12 [ CrossRef ] [ Google Scholar ]
- Berlingeri M., Bottini G., Basilico S., Silani G., Zanardi G., Sberna M., et al. (2008). Anatomy of the episodic buffer: a Voxel-based morphometry study in patients with dementia. Behav. Neurol. 19 29–34. 10.1155/2008/828937 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Bermudez-Rattoni F. (2010). Is memory consolidation a multiple-circuit system? Proc. Natl. Acad. Sci. U.S.A. 107 8051–8052. 10.1073/pnas.1003434107 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Binder J. R., Desai R. H. (2011). The neurobiology of semantic memory. Trends Cogn. Sci. 15 527–536. 10.1016/j.tics.2011.10.001 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Binney R. J., Hoffman P., Lambon Ralph M. A. (2016). Mapping the multiple graded contributions of the anterior temporal lobe representational hub to abstract and social concepts: evidence from distortion-corrected fmri. Cereb. Cortex 26 4227–4241. 10.1093/cercor/bhw260 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Bliss T. V., Collingridge G. L. (1993). A synaptic model of memory: long-term potentiation in the hippocampus. Nature 361 31–39. 10.1038/361031a0 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Blumenfeld R. S., Ranganath C. (2007). Prefrontal cortex and long-term memory encoding: an integrative review of findings from Neuropsychology and Neuroimaging. Neuroscientist 13 280–291. 10.1177/1073858407299290 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Boran E., Fedele T., Klaver P., Hilfiker P., Stieglitz L., Grunwald T., et al. (2019). Persistent hippocampal neural firing and hippocampal-cortical coupling predict verbal working memory load. Sci. Adv. 5 : eaav3687 . 10.1126/sciadv.aav3687 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Born J., Rasch B., Gais S. (2006). Sleep to remember. Neuroscientist 12 410–424. 10.1177/1073858406292647 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Braver T. S., Gray J. R., Burgess G. C. (2008). “ Explaining the many varieties of working memory variation: dual mechanisms of cognitive control ,” in Variation in working memory , 1st Edn, eds Conway A., Jarrold C., Kane M., Miyake A., Towse J. (New York, NY: Oxford University Press; ), 76–106. 10.1093/acprof:oso/9780195168648.003.0004 [ CrossRef ] [ Google Scholar ]
- Brown T. I., Rissman J., Chow T. E., Uncapher M. R., Wagner A. D. (2018). Differential medial temporal lobe and parietal cortical contributions to real-world autobiographical episodic and autobiographical semantic memory. Sci. Rep. 8 : 6190 . 10.1038/s41598-018-24549-y [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Buchsbaum B. R. (2010). Neural basis of working memory. Encyclopedia of Behavioral Neuroscience 334–341. 10.1016/b978-0-08-045396-5.00161-5 [ CrossRef ] [ Google Scholar ]
- Buchsbaum B. R., Hickok G., Humphries C. (2001). Role of left posterior superior temporal gyrus in phonological processing for speech perception and production. Cogn. Sci. 25 663–678. 10.1207/s15516709cog2505_2 [ CrossRef ] [ Google Scholar ]
- Buiatti T., Mussoni A., Toraldo A., Skrap M., Shallice T. (2011). Two qualitatively different impairments in making rotation operations. Cortex 47 166–179. 10.1016/j.cortex.2009.10.006 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Burbaud P., Camus O., Caillé J., Biolac B., Allard M. (1999). Influence of individual strategies on brain activation patterns during cognitive tasks. J. Neuroradiol. 26 59–65. [ PubMed ] [ Google Scholar ]
- Camina E., Güell F. (2017). The neuroanatomical, neurophysiological and psychological basis of memory: current models and their origins. Front. Pharmacol. 8 : 438 . 10.3389/fphar.2017.00438 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Caplan D., Waters G. (2005). The relationship between age, processing speed, working memory capacity, and language comprehension. Memory 13 403–413. 10.1080/09658210344000459 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Chein J. M., Ravizza S. M., Fiez J. A. (2003). Using neuroimaging to evaluate models of working memory and their implications for language processing. J. Neurolinguist. 16 315–339. 10.1016/s0911-6044(03)00021-6 [ CrossRef ] [ Google Scholar ]
- Clawson B. C., Pickup E. J., Ensing A., Geneseo L., Shaver J., Gonzalez-Amoretti J., et al. (2021). Causal role for sleep-dependent reactivation of learning-activated sensory ensembles for fear memory consolidation . Nat. Commun . 12 , 1–13. 10.1038/s41467-021-21471-2 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Cohen M. S., Kosslyn S. M., Breiter H. C., DiGirolamo G. J., Thompson W. L., Anderson A. K., et al. (1996). Changes in cortical activity during mental rotation a mapping study using functional MRI. Brain 119 89–100. 10.1093/brain/119.1.89 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Cooper R. A., Ritchey M. (2019). Progression from feature-specific brain activity to hippocampal binding during episodic encoding. J. Neurosci. 40 1701–1709. 10.1523/jneurosci.1971-19.2019 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Curtis C. E., Sprague T. C. (2021). Persistent activity during working memory from front to back. Front. Neural Circ. 15 : 696060 . 10.3389/fncir.2021.696060 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- D’Esposito M. (2007). From cognitive to neural models of working memory. Philos. Trans. R. Soc. B Biol. Sci. 362 761–772. 10.1098/rstb.2007.2086 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- D’Esposito M., Postle B. R. (2015). The cognitive neuroscience of working memory. Annu. Rev. Psychol. 66 115–142. 10.1146/annurev-psych-010814-015031 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Daselaar S. M. (2009). Posterior midline and ventral parietal activity is associated with retrieval success and encoding failure. Front. Hum. Neurosci. 3 : 13 . 10.3389/neuro.09.013.2009 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Datta S., MacLean R. R. (2007). Neurobiological mechanisms for the regulation of mammalian sleep–wake behavior: reinterpretation of historical evidence and inclusion of contemporary cellular and molecular evidence. Neurosci. Biobehav. Rev. 31 775–824. 10.1016/j.neubiorev.2007.02.004 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Davis S. W., Wing E. A., Cabeza R. (2018). Contributions of the ventral parietal cortex to declarative memory. Handb. Clin. Neurol. 151 525–553. 10.1016/b978-0-444-63622-5.00027-9 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Derner M., Dehnen G., Chaieb L., Reber T. P., Borger V., Surges R., et al. (2020). Patterns of single-neuron activity during associative recognition memory in the human medial temporal lobe. Neuroimage 221 : 117214 . 10.1016/j.neuroimage.2020.117214 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Dew I. T. Z., Cabeza R. (2011). The porous boundaries between explicit and implicit memory: behavioral and neural evidence. Ann. N. Y. Acad. Sci. 1224 174–190. 10.1111/j.1749-6632.2010.05946.x [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Diekelmann S., Born J. (2010). The memory function of sleep. Nat. Rev. Neurosci. 11 114–126. 10.1038/nrn2762 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Dudai Y. (2002). Molecular bases of long-term memories: a question of persistence. Curr. Opin. Neurobiol. 12 211–216. 10.1016/s0959-4388(02)00305-7 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Dudai Y., Karni A., Born J. (2015). The consolidation and transformation of memory. Neuron 88 20–32. 10.1016/j.neuron.2015.09.004 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Dunsmoor J. E., Murty V. P., Davachi L., Phelps E. A. (2015). Emotional learning selectively and retroactively strengthens memories for related events. Nature 520 345–348. 10.1038/nature14106 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Düzel E., Penny W. D., Burgess N. (2010). Brain oscillations and memory. Curr. Opin. Neurobiol. 20 143–149. 10.1016/j.conb.2010.01.004 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Eichenbaum H., Cohen N. J. (2014). Can we reconcile the declarative memory and spatial navigation views on hippocampal function? Neuron 83 764–770. 10.1016/j.neuron.2014.07.032 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Eichenlaub J.-B., Jarosiewicz B., Saab J., Franco B., Kelemen J., Halgren E., et al. (2020). Replay of learned neural firing sequences during rest in human motor cortex. Cell Rep. 31 : 107581 . 10.1016/j.celrep.2020.107581 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Emch M., von Bastian C. C., Koch K. (2019). Neural correlates of verbal working memory: an fMRI meta-analysis. Front. Hum. Neurosci. 13 : 180 . 10.3389/fnhum.2019.00180 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Frankland P. W., Bontempi B. (2005). The organization of recent and remote memories. Nature Rev. Neurosci. 6 119–130. 10.1038/nrn1607 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Frey U., Morris R. G. M. (1998). Synaptic tagging: implications for late maintenance of hippocampal long-term potentiation. Trends Neurosci. 21 181–188. 10.1016/s0166-2236(97)01189-2 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Friedman N. P., Robbins T. W. (2022). The role of prefrontal cortex in cognitive control and executive function. Neuropsychopharmacology 47 72–89. 10.1038/s41386-021-01132-0 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Funahashi S. (2017). Working memory in the prefrontal cortex. Brain Sci. 7 : 49 . 10.3390/brainsci7050049 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Gais S., Lucas B., Born J. (2006). Sleep after learning AIDS memory recall. Learn. Mem. 13 259–262. 10.1101/lm.132106 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Gallistel C. R., Balsam P. D. (2014). Time to rethink the neural mechanisms of learning and memory. Neurobiol. Learn. Mem. 108 136–144. 10.1016/j.nlm.2013.11.019 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Ge Y., Dong Z., Bagot R. C., Howland J. G., Phillips A. G., Wong T. P., et al. (2010). Hippocampal long-term depression is required for the consolidation of Spatial Memory. Proc. Natl Acad. Sci. U.S.A. 107 16697–16702. 10.1073/pnas.1008200107 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Gelastopoulos A., Whittington M. A., Kopell N. J. (2019). Parietal low beta rhythm provides a dynamical substrate for a working memory buffer. Proc. Natl Acad. Sci. U.S.A. 116 16613–16620. 10.1073/pnas.1902305116 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Genzel L., Wixted J. T. (2017). “ Cellular and systems consolidation of declarative memory ,” in Cognitive neuroscience of memory consolidation , eds Axmacher N., Rasch B. (Berlin: Springer International Publishing; ), 3–16. 10.1007/978-3-319-45066-7_1 [ CrossRef ] [ Google Scholar ]
- Ghaleh M., Lacey E. H., Fama M. E., Anbari Z., DeMarco A. T., Turkeltaub P. E. (2019). Dissociable mechanisms of verbal working memory revealed through multivariate lesion mapping. Cereb. Cortex 30 2542–2554. 10.1093/cercor/bhz259 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Gilboa A., Moscovitch M. (2021). No consolidation without representation: correspondence between neural and psychological representations in recent and remote memory. Neuron 109 2239–2255. 10.1016/j.neuron.2021.04.025 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Girardeau G., Lopes-dos-Santos V. (2021). Brain neural patterns and the memory function of sleep. Science 374 560–564. 10.1126/science.abi8370 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Goedert K. M., Willingham D. B. (2002). Patterns of interference in sequence learning and prism adaptation inconsistent with the consolidation hypothesis. Learn. Mem. 9 279–292. 10.1101/lm.50102 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Goerke M., Cohrs S., Rodenbeck A., Grittner U., Sommer W., Kunz D. (2013). Declarative memory consolidation during the first night in a sleep lab: the role of REM sleep and Cortisol. Psychoneuroendocrinology 38 1102–1111. 10.1016/j.psyneuen.2012.10.019 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Graves W. W., Grabowski T. J., Mehta S., Gupta P. (2008). The left posterior superior temporal gyrus participates specifically in accessing lexical phonology. J. Cogn. Neurosci. 20 1698–1710. 10.1162/jocn.2008.20113 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Hamann S. (2001). Cognitive and neural mechanisms of emotional memory. Trends Cogn. Sci. 5 394–400. 10.1016/S1364-6613(00)01707-1 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Henson R. N., Rugg M. D., Shallice T., Josephs O., Dolan R. J. (1999). Recollection and familiarity in recognition memory: an event-related functional magnetic resonance imaging study. J. Neurosci. 19 3962–3972. 10.1523/jneurosci.19-10-03962.1999 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Herry C., Johansen J. P. (2014). Encoding of fear learning and memory in distributed neuronal circuits. Nat. Neurosci. 17 1644–1654. 10.1038/nn.3869 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Hitch G. J., Allen R. J., Baddeley A. D. (2019). Attention and binding in visual working memory: two forms of attention and two kinds of buffer storage. Attent. Percept. Psychophys. 82 280–293. 10.3758/s13414-019-01837-x [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Howard M. D., Skorheim S. W., Pilly P. K. (2022). A model of bi-directional interactions between complementary learning systems for memory consolidation of sequential experiences. Front. Syst. Neurosci. 16 : 972235 . 10.3389/fnsys.2022.972235 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Hutchinson J. B., Uncapher M. R., Wagner A. D. (2009). Posterior parietal cortex and episodic retrieval: convergent and divergent effects of attention and memory. Learn. Mem. 16 343–356. 10.1101/lm.919109 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Ikkai A., Curtis C. E. (2011). Common neural mechanisms supporting spatial working memory, attention and motor intention. Neuropsychologia 49 1428–1434. 10.1016/j.neuropsychologia.2010.12.020 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Ito M., Kuroiwa J., Miyake S. (2003). A neural network model of memory system using hippocampus. Electron. Commun. Japan 86 86–97. 10.1002/ecjc.1010 [ CrossRef ] [ Google Scholar ]
- Ivanco T. L. (2015). Long-term potentiation and long-term depression. International Encyclopedia of the Social & Behavioral Sciences 14 , 358–365. 10.1016/b978-0-08-097086-8.55034-x [ CrossRef ] [ Google Scholar ]
- Jerde T. A., Curtis C. E. (2013). Maps of space in human frontoparietal cortex. J. Physiol. 107 510–516. 10.1016/j.jphysparis.2013.04.002 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Ji D., Wilson M. A. (2006). Coordinated memory replay in the visual cortex and hippocampus during sleep. Nat. Neurosci. 10 100–107. 10.1038/nn1825 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Josselyn S. A., Köhler S., Frankland P. W. (2015). Finding the engram. Nat. Rev. Neurosci. 16 521–534. 10.1038/nrn4000 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Jung J., Lambon Ralph M. A., Jackson R. L. (2022). Subregions of DLPFC display graded yet distinct structural and functional connectivity. J. Neurosci. 42 3241–3252. 10.1523/jneurosci.1216-21.2022 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Kaleem A., Hoessli D. C., Haq I., Walker-Nasir E., Butt A., Iqbal Z., et al. (2011). CREB in long-term potentiation in hippocampus: role of post-translational modifications-studies in silico. J. Cell. Biochem. 112 138–146. 10.1002/jcb.22909 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Kamiński J., Rutishauser U. (2019). Between persistently active and activity-silent frameworks: novel vistas on the cellular basis of working memory. Ann. N. Y. Acad. Sci. 1464 64–75. 10.1111/nyas.14213 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Kamiński J., Sullivan S., Chung J. M., Ross I. B., Mamelak A. N., Rutishauser U. (2017). Persistently active neurons in human medial frontal and medial temporal lobe support working memory. Nat. Neurosci. 20 590–601. 10.1038/nn.4509 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Kandel E. R., Mack S. (2013). Principles of neural science. New York, NY: McGraw-Hill Medical. [ Google Scholar ]
- Kandel E. R., Dudai Y., Mayford M. R. (2014). The molecular and systems biology of memory. Cell 157 163–186. 10.1016/j.cell.2014.03.001 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Karni A., Meyer G., Rey-Hipolito C., Jezzard P., Adams M. M., Turner R., et al. (1998). The acquisition of skilled motor performance: fast and slow experience-driven changes in primary motor cortex. Proc. Natl. Acad. Sci. U.S.A. 95 861–868. 10.1073/pnas.95.3.861 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Kerzel D., Burra N. (2020). Capture by context elements, not attentional suppression of distractors, explains the PD with small search displays. J. Cogn. Neurosci. 32 1170–1183. 10.1162/jocn_a_01535 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Kesner R. P. (2017). Memory neurobiology?. Reference Module in Neuroscience and Biobehavioral Psychology 1–12. 10.1016/b978-0-12-809324-5.03089-3 [ CrossRef ] [ Google Scholar ]
- Kim J., Gulati T., Ganguly K. (2019). Competing roles of slow oscillations and delta waves in memory consolidation versus forgetting. Cell 179 514–526.e13. 10.1016/j.cell.2019.08.040 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Koenigs M., Acheson D. J., Barbey A. K., Solomon J., Postle B. R., Grafman J. (2011). Areas of left perisylvian cortex mediate auditory–verbal short-term memory. Neuropsychologia 49 3612–3619. 10.1016/j.neuropsychologia.2011.09.013 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Kosslyn S. M., Thompson W. L., Alpert N. M. (1997). Neural Systems shared by visual imagery and visual perception: a positron emission tomography study. Neuroimage 6 320–334. 10.1006/nimg.1997.0295 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Kuhnke P., Kiefer M., Hartwigsen G. (2020). Task-dependent recruitment of modality-specific and multimodal regions during conceptual processing. Cereb. Cortex 30 3938–3959. 10.1093/cercor/bhaa010 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Landmann N., Kuhn M., Piosczyk H., Feige B., Baglioni C., Spiegelhalder K., et al. (2014). The reorganisation of memory during sleep. Sleep Med. Rev. 18 531–541. 10.1016/j.smrv.2014.03.005 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Langille J. J. (2019). Remembering to forget: a dual role for sleep oscillations in memory consolidation and forgetting. Front. Cell. Neurosci. 13 : 71 . 10.3389/fncel.2019.00071 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Langille J. J., Brown R. E. (2018). The synaptic theory of memory: a historical survey and reconciliation of recent opposition. Front. Syst. Neurosci. 12 : 52 . 10.3389/fnsys.2018.00052 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Lee H., Chun M. M., Kuhl B. A. (2016). Lower parietal encoding activation is associated with sharper information and Better Memory. Cereb. Cortex 27 , 2486–2499. 10.1093/cercor/bhw097 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Li M., Lu S., Wu Y., Zhong N. (2010). “ Functional segregation of memory encoding and retrieval in the prefrontal cortex ,” in Proceedings of the IEEE/ICME International Conference on Complex Medical Engineering , (Gold Coast, QLD: ), 91–95. 10.1109/iccme.2010.5558865 [ CrossRef ] [ Google Scholar ]
- Li Q., Song J.-L., Li S.-H., Westover M. B., Zhang R. (2020). Effects of cholinergic neuromodulation on thalamocortical rhythms during NREM sleep: a model study. Front. Comput. Neurosci. 13 : 100 . 10.3389/fncom.2019.00100 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Lisman J. E., Grace A. A. (2005). The hippocampal-VTA loop: controlling the entry of information into long-term memory. Neuron 46 703–713. 10.1016/j.neuron.2005.05.002 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Liu J., Zhang H., Yu T., Ren L., Ni D., Yang Q., et al. (2021). Transformative neural representations support long-term episodic memory. Sci. Adv. 7 : eabg9715 . 10.1126/sciadv.abg9715 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Luscher C., Malenka R. C. (2012). NMDA receptor-dependent long-term potentiation and long-term depression (LTP/LTD). Cold Spring Harb. Perspect. Biol. 4 a005710–a005710. 10.1101/cshperspect.a005710 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Lynch M. A. (2004). Long-term potentiation and memory. Physiol. Rev. 84 87–136. 10.1152/physrev.00014.2003 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Maass W., Markram H. (2002). Synapses as dynamic memory buffers. Neural Netw. 15 155–161. 10.1016/s0893-6080(01)00144-7 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Mackey W. E., Devinsky O., Doyle W. K., Golfinos J. G., Curtis C. E. (2016). Human parietal cortex lesions impact the precision of spatial working memory. J. Neurophysiol. 116 1049–1054. 10.1152/jn.00380.2016 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Malenka R. C., Bear M. F. (2004). LTP and Ltd. Neuron 44 5–21. 10.1016/j.neuron.2004.09.012 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Malenka R. C., Nicoll R. (1999). Long-term potentiation–a decade of progress? Science 285 1870–1874. 10.1126/science.285.5435.1870 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Mank M., Griesbeck O. (2008). Genetically encoded calcium indicators. Chem. Rev. 108 1550–1564. 10.1021/cr078213v [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Maquet P. (2001). The role of sleep in learning and memory. Science 294 1048–1052. 10.1126/science.1062856 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Marshall L., Born J. (2007). The contribution of sleep to hippocampus-dependent memory consolidation. Trends Cogn. Sci. 11 442–450. 10.1016/j.tics.2007.09.001 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Mayes A. R. (2002). Memory disorders, organic. Encyclopedia of the Human Brain 2 , 759–772. 10.1016/b0-12-227210-2/00199-0 [ CrossRef ] [ Google Scholar ]
- McGaugh J. L. (2000). Memory–a century of consolidation. Science 287 248–251. 10.1126/science.287.5451.248 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Mednick S. C., Cai D. J., Shuman T., Anagnostaras S., Wixted J. T. (2011). An opportunistic theory of cellular and systems consolidation. Trends Neurosci. 34 504–514. 10.1016/j.tins.2011.06.003 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Menon V., D’Esposito M. (2021). The role of PFC networks in cognitive control and executive function. Neuropsychopharmacology 47 90–103. 10.1038/s41386-021-01152-w [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Miyake A., Friedman N. P., Rettinger D. A., Shah P., Hegarty M. (2001). How are visuospatial working memory, executive functioning, and spatial abilities related? A latent-variable analysis. J. Exp. Psychol. 130 621–640. 10.1037/0096-3445.130.4.621 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Moncada D., Ballarini F., Viola H. (2015). Behavioral tagging: a translation of the synaptic tagging and capture hypothesis. Neural Plast. 2015 1–21. 10.1155/2015/650780 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Moro V., Berlucchi G., Lerch J., Tomaiuolo F., Aglioti S. M. (2008). Selective deficit of mental visual imagery with intact primary visual cortex and visual perception. Cortex 44 109–118. 10.1016/j.cortex.2006.06.004 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Moscovitch M., Nadel L., Winocur G., Gilboa A., Rosenbaum R. S. (2006). The cognitive neuroscience of remote episodic, semantic and spatial memory. Curr. Opin. Neurobiol. 16 179–190. 10.1016/j.conb.2006.03.013 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Nabavi S., Fox R., Proulx C. D., Lin J. Y., Tsien R. Y., Malinow R. (2014). Engineering A memory with ltd and LTP. Nature 511 348–352. 10.1038/nature13294 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Nader K., Hardt O. (2009). A single standard for memory: the case for reconsolidation. Nat. Rev. Neurosci. 10 224–234. 10.1038/nrn2590 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Narayanan N. S., Prabhakaran V., Bunge S. A., Christoff K., Fine E. M., Gabrieli J. D. E. (2005). The role of the prefrontal cortex in the maintenance of verbal working memory: an event-related fMRI analysis. Neuropsychology 19 223–232. 10.1037/0894-4105.19.2.223 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Nee D. E., D’Esposito M. (2016). The hierarchical organization of the lateral prefrontal cortex. eLife 5 : e12112 . 10.7554/elife.12112 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Ngo H.-V., Fell J., Staresina B. (2020). Sleep spindles mediate hippocampal-neocortical coupling during long-duration ripples. eLife 9 : e57011 . 10.7554/elife.57011 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Ober B. A. (2014). Memory, explicit/implicit. Encyclopedia of the Neurological Sciences 2 , 1042–1044. 10.1016/b978-0-12-385157-4.00455-3 [ CrossRef ] [ Google Scholar ]
- Okuda K., Højgaard K., Privitera L., Bayraktar G., Takeuchi T. (2020). Initial memory consolidation and the synaptic tagging and capture hypothesis. Eur. J. Neurosci. 54 6826–6849. 10.1111/ejn.14902 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Orsini C. A., Maren S. (2012). Neural and cellular mechanisms of fear and extinction memory formation. Neurosci. Biobehav. Rev. 36 1773–1802. 10.1016/j.neubiorev.2011.12.014 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Osaka N., Logie R. H., D’Esposito M. (eds) (2007). The cognitive neuroscience of working memory. Oxford: Oxford University Press. [ Google Scholar ]
- Panzeri S., Janotte E., Pequeńo-Zurro A., Bonato J., Bartolozzi C. (2023). Constraints on the design of neuromorphic circuits set by the properties of neural population codes. Neuromorphic Computing Eng. 3 : 012001 . 10.1088/2634-4386/acaf9c [ CrossRef ] [ Google Scholar ]
- Park P., Kang H., Georgiou J., Zhuo M., Kaang B.-K., Collingridge G. L. (2021). Further evidence that CP-ampars are critically involved in synaptic tag and capture at hippocampal CA1 synapses. Mol. Brain 14 : 26 . 10.1186/s13041-021-00737-2 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Park P., Kang H., Sanderson T. M., Bortolotto Z. A., Georgiou J., Zhuo M., et al. (2018). The role of calcium-permeable ampars in long-term potentiation at principal neurons in the rodent hippocampus. Front. Synaptic Neurosci. 10 : 42 . 10.3389/fnsyn.2018.00042 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Patterson K., Nestor P. J., Rogers T. T. (2007). Where do you know what you know? The representation of semantic knowledge in the human brain. Nat. Rev. Neurosci. 8 976–987. 10.1038/nrn2277 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Paulesu E., Frith C. D., Frackowiak R. S. (1993). The neural correlates of the verbal component of working memory. Nature 362 342–345. 10.1038/362342a0 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Perrachione T. K., Ghosh S. S., Ostrovskaya I., Gabrieli J. D., Kovelman I. (2017). Phonological working memory for words and nonwords in cerebral cortex. J. Speech Lang. Hear. Res. 60 1959–1979. 10.1044/2017_jslhr-l-15-0446 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Popa D., Duvarci S., Popescu A. T., Léna C., Paré D. (2010). Coherent amygdalocortical theta promotes fear memory consolidation during paradoxical sleep. Proc. Natl. Acad. Sci. U.S.A. 107 6516–6519. 10.1073/pnas.0913016107 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Postle B. R. (2006). Working memory as an emergent property of the mind and brain. Neuroscience 139 23–38. 10.1016/j.neuroscience.2005.06.005 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Postle B. R. (2015). The cognitive neuroscience of visual short-term memory. Curr. Opin. Behav. Sci. 1 40–46. 10.1016/j.cobeha.2014.08.004 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Puentes-Mestril C., Aton S. J. (2017). Linking network activity to synaptic plasticity during sleep: hypotheses and recent data. Front. Neural Circ. 11 : 61 . 10.3389/fncir.2017.00061 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Qasim S. E., Mohan U. R., Stein J. M., Jacobs J. (2023). Neuronal activity in the human amygdala and hippocampus enhances emotional memory encoding. Nat. Hum. Behav. 7 754–764. 10.1038/s41562-022-01502-8 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Quentin R., King J.-R., Sallard E., Fishman N., Thompson R., Buch E. R., et al. (2019). Differential brain mechanisms of selection and maintenance of information during working memory. J. Neurosci. 39 3728–3740. 10.1523/jneurosci.2764-18.2019 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Rakowska M., Bagrowska P., Lazari A., Navarrete M., Abdellahi M. E., Johansen-Berg H., et al. (2022). Cueing motor memory reactivation during NREM sleep engenders learning-related changes in precuneus and sensorimotor structures . bioRxiv 2022.01.-27.477838. 10.1101/2022.01.27.477838 [ CrossRef ] [ Google Scholar ]
- Rasch B., Born J. (2007). Maintaining memories by reactivation. Curr. Opin. Neurobiol. 17 698–703. 10.1016/j.conb.2007.11.007 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Rasch B., Born J. (2013). About sleep’s role in memory. Physiol. Rev. 93 681–766. 10.1152/physrev.00032.2012 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Ratcliffe O., Shapiro K., Staresina B. P. (2022). Fronto-medial theta coordinates posterior maintenance of working memory content. Curr. Biol. 32 2121–2129.e3. 10.1016/j.cub.2022.03.045 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Reber P. J. (2008). Cognitive neuroscience of declarative and nondeclarative memory. Hum. Learn. Biol. Brain Neurosci. 139 113–123. 10.1016/s0166-4115(08)10010-3 [ CrossRef ] [ Google Scholar ]
- Redondo R. L., Morris R. G. M. (2011). Making memories last: the synaptic tagging and capture hypothesis. Nat. Rev. Neurosci. 12 17–30. 10.1038/nrn2963 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Ren Z., Zhang Y., He H., Feng Q., Bi T., Qiu J. (2019). The different brain mechanisms of object and spatial working memory: voxel-based morphometry and resting-state functional connectivity. Front. Hum. Neurosci. 13 : 248 . 10.3389/fnhum.2019.00248 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Repovs G., Baddeley A. (2006). The multi-component model of working memory: explorations in experimental cognitive psychology. Neuroscience 139 5–21. 10.1016/j.neuroscience.2005.12.061 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Reyes-Resina I., Samer S., Kreutz M. R., Oelschlegel A. M. (2021). Molecular mechanisms of memory consolidation that operate during sleep. Front. Mol. Neurosci. 14 : 767384 . 10.3389/fnmol.2021.767384 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Ribeiro S. (2007). Novel experience induces persistent sleep-dependent plasticity in the cortex but not in the hippocampus. Front. Neurosci. 1 :43–55. 10.3389/neuro.01.1.1.003.2007 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Richter-Levin G., Akirav I. (2003). Emotional tagging of memory formation—In the search for neural mechanisms. Brain Res. Rev. 43 247–256. 10.1016/j.brainresrev.2003.08.005 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Ritchey M., Dolcos F., Cabeza R. (2008). Role of amygdala connectivity in the persistence of emotional memories over time: an event-related fmri investigation. Cereb. Cortex 18 2494–2504. 10.1093/cercor/bhm262 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Robin J., Moscovitch M. (2017). Details, gist and schema: hippocampal–neocortical interactions underlying recent and remote episodic and spatial memory. Curr. Opin. Behav. Sci. 17 114–123. 10.1016/j.cobeha.2017.07.016 [ CrossRef ] [ Google Scholar ]
- Robins S. K. (2020). Stable engrams and neural dynamics. Philos. Sci. 87 1130–1139. 10.1086/710624 [ CrossRef ] [ Google Scholar ]
- Roediger H. L., Butler A. C. (2011). The critical role of retrieval practice in long-term retention. Trends Cogn. Sci. 15 20–27. 10.1016/j.tics.2010.09.003 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Roldan-Valadez E., García-Lázaro H., Lara-Romero R., Ramirez-Carmona R. (2012). Neuroanatomy of episodic and semantic memory in humans: a brief review of neuroimaging studies. Neurol. India 60 : 613 . 10.4103/0028-3886.105196 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Saffran N. M. (1997). Language and auditory-verbal short-term memory impairments: evidence for common underlying processes. Cogn. Neuropsychol. 14 641–682. 10.1080/026432997381402 [ CrossRef ] [ Google Scholar ]
- Salvato G., Peviani V., Scarpa P., Francione S., Castana L., Gallace A., et al. (2021). Investigating visuo-spatial neglect and visual extinction during intracranial electrical stimulations: the role of the right inferior parietal cortex. Neuropsychologia 162 : 108049 . 10.1016/j.neuropsychologia.2021.108049 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Sarnthein J., Petsche H., Rappelsberger P., Shaw G. L., von Stein A. (1998). Synchronization between prefrontal and posterior association cortex during human working memory. Proc. Natl. Acad. Sci. U.S.A. 95 7092–7096. 10.1073/pnas.95.12.7092 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Schiller D., Monfils M.-H., Raio C. M., Johnson D. C., LeDoux J. E., Phelps E. A. (2009). Preventing the return of fear in humans using reconsolidation update mechanisms. Nature 463 49–53. 10.1038/nature08637 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Schlichting M. L., Preston A. R. (2015). Memory integration: neural mechanisms and implications for behavior. Curr. Opin. Behav. Sci. 1 1–8. 10.1016/j.cobeha.2014 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Sellers K. K., Yu C., Zhou Z. C., Stitt I., Li Y., Radtke-Schuller S., et al. (2016). Oscillatory dynamics in the frontoparietal attention network during sustained attention in the ferret. Cell Rep. 16 2864–2874. 10.1016/j.celrep.2016.08.055 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Sendi M. S., Kanta V., Inman C. S., Manns J. R., Hamann S., Gross R. E., et al. (2020). “ Amygdala stimulation leads to functional network connectivity state transitions in the hippocampus ,” in Proceedings of the 2020 42nd Annual International Conference of the IEEE Engineering in Medicine & Biology Society (EMBC) , (Montreal, QC: ). 10.1109/embc44109.2020.9176742 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Serences J. T. (2016). Neural mechanisms of information storage in visual short-term memory. Vis. Res. 128 53–67. 10.1016/j.visres.2016.09.010 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Shafritz K. M., Gore J. C., Marois R. (2002). The role of the parietal cortex in visual feature binding. Proc. Natl. Acad. Sci. U.S.A. 99 10917–10922. 10.1073/pnas.152694799 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Shallice T. (2002). “ Fractionation of the supervisory system ,” in Principles of frontal lobe function , eds Stuss D. T., Knight R. T. (Oxford: Oxford University Press; ), 261–277. 10.1093/acprof:oso/9780195134971.003.0017 [ CrossRef ] [ Google Scholar ]
- Siegel J. M. (2001). The REM sleep-memory consolidation hypothesis. Science 294 1058–1063. 10.1126/science.1063049 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Smith E. E., Jonides J., Marshuetz C., Koeppe R. A. (1998). Components of verbal working memory: evidence from neuroimaging. Proc. Natl. Acad. Sci. U.S.A. 95 876–882. 10.1073/pnas.95.3.876 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Sobczak J. (2017). Mechanisms of memory consolidation (dissertation). PhD thesis. York: University of York. [ Google Scholar ]
- Sorensen K. E. (2009). The connections of the hippocampal region new observations on efferent connections in the guinea pig, and their functional implications*. Acta Neurol. Scand. 72 550–560. 10.1111/j.1600-0404.1985.tb00914.x [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Speranza L., di Porzio U., Viggiano D., de Donato A., Volpicelli F. (2021). Dopamine: the neuromodulator of long-term synaptic plasticity, reward and movement control. Cells 10 : 735 . 10.3390/cells10040735 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Squire L. R., Wixted J. T. (2011). The cognitive neuroscience of human memory since H.M. Annu. Rev. Neurosci. 34 259–288. 10.1146/annurev-neuro-061010-113720 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Squire L. R., Zola S. M. (1996). Structure and function of declarative and nondeclarative memory systems. Proc. Natl. Acad. Sci. U.S.A. 93 13515–13522. 10.1073/pnas.93.24.13515 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Squire L. R., Genzel L., Wixted J. T., Morris R. G. (2015). Memory consolidation. Cold Spring Harb. Perspect. Biol. 7 : a021766 . 10.1101/cshperspect.a021766 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Squire L. R., Stark C. E. L., Clark R. E. (2004). The medial temporal lobe. Annu. Rev. Neurosci. 27 279–306. 10.1146/annurev.neuro.27.070203.144130 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Sreenivasan K. K., Curtis C. E., D’Esposito M. (2014). Revisiting the role of persistent neural activity during working memory. Trends Cogn. Sci. 18 82–89. 10.1016/j.tics.2013.12.001 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Stickgold R. (2005). Sleep-dependent memory consolidation. Nature 437 1272–1278. 10.1038/nature04286 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Swanson R. A., Levenstein D., McClain K., Tingley D., Buzsáki G. (2020). Variable specificity of memory trace reactivation during hippocampal sharp wave ripples. Curr. Opin. Behav. Sci. 32 126–135. 10.1016/j.cobeha.2020.02.008 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Takehara-Nishiuchi K. (2020). Prefrontal–hippocampal interaction during the encoding of New Memories. Brain Neurosci. Adv. 4 239821282092558 . 10.1177/2398212820925580 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Takeuchi T., Duszkiewicz A. J., Morris R. G. (2014). The synaptic plasticity and memory hypothesis: encoding, storage and persistence. Philos. Trans. R. Soc. B Biol. Sci. 369 : 20130288 . 10.1098/rstb.2013.0288 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Tamminen J., Payne J. D., Stickgold R., Wamsley E. J., Gaskell M. G. (2010). Sleep spindle activity is associated with the integration of new memories and existing knowledge. The Journal of Neuroscience 30 14356–14360. 10.1523/jneurosci.3028-10.2010 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Thomas Yeo B. T., Krienen F. M., Sepulcre J., Sabuncu M. R., Lashkari D., Hollinshead M., et al. (2011). The organization of the human cerebral cortex estimated by intrinsic functional connectivity. J. Neurophysiol. 106 1125–1165. 10.1152/jn.00338.2011 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Thompson R. F., Kim J. J. (1996). Memory systems in the brain and localization of a memory. Proc. Natl. Acad. Sci. U.S.A. 93 13438–13444. 10.1073/pnas.93.24.13438 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Thuault S. J., Malleret G., Constantinople C. M., Nicholls R., Chen I., Zhu J., et al. (2013). Prefrontal cortex HCN1 channels enable intrinsic persistent neural firing and executive memory function. J. Neurosci. 33 13583–13599. 10.1523/jneurosci.2427-12.2013 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Todd J. J., Marois R. (2005). Posterior parietal cortex activity predicts individual differences in visual short-term memory capacity. Cogn. Affect. Behav. Neurosci. 5 144–155. 10.3758/cabn.5.2.144 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Tononi G., Cirelli C. (2003). Sleep and synaptic homeostasis: a hypothesis. Brain Res. Bull. 62 143–150. 10.1016/j.brainresbull.2003.09.004 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Tononi G., Cirelli C. (2014). Sleep and the price of plasticity: from synaptic and cellular homeostasis to memory consolidation and integration. Neuron 81 12–34. 10.1016/j.neuron.2013.12.025 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Tulving E. (1985). Memory and consciousness. Can. Psychol. 26 1–12. 10.1037/h0080017 [ CrossRef ] [ Google Scholar ]
- van Kesteren M. T. R., Ruiter D. J., Fernández G., Henson R. N. (2012). How schema and novelty augment memory formation. Trends Neurosci. 35 211–219. 10.1016/j.tins.2012.02.001 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- van Kesteren M. T., Brown T. I., Wagner A. D. (2016). Interactions between memory and new learning: insights from fMRI multivoxel pattern analysis. Front. Syst. Neurosci. 10 : 46 . 10.3389/fnsys.2016.00046 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Vilberg K. L., Rugg M. D. (2008). Memory retrieval and the PARIETAL CORTEX: a review of evidence from a dual-process perspective. Neuropsychologia 46 1787–1799. 10.1016/j.neuropsychologia.2008.01.004 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Vishnoi S., Naseem M., Raisuddin S., Parvez S. (2018). Behavioral tagging: plausible involvement of PKMZ, ARC and role of neurotransmitter receptor systems. Neurosci. Biobehav. Rev. 94 210–218. 10.1016/j.neubiorev.2018.07.009 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Wang J. H., Wang D., Gao Z., Chen N., Lei Z., Cui S., et al. (2016). Both glutamatergic and Gabaergic neurons are recruited to be associative memory cells. Biophys. J. 110 : 481a . 10.1016/j.bpj.2015.11.2571 [ CrossRef ] [ Google Scholar ]
- Wang J.-H., Cui S. (2018). Associative memory cells and their working principle in the brain. F1000Research 7 : 108 . 10.12688/f1000research.13665.1 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Wig G. S., Buckner R. L., Schacter D. L. (2009). Repetition priming influences distinct brain systems: evidence from task-evoked data and resting-state correlations. J. Neurophysiol. 101 2632–2648. 10.1152/jn.91213.2008 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Williams J. N. (2020). The neuroscience of implicit learning. Language Learning 70 255–307. 10.1111/lang.12405 [ CrossRef ] [ Google Scholar ]
- Wiltgen B. J., Zhou M., Cai Y., Balaji J., Karlsson M. G., Parivash S. N., et al. (2010). The hippocampus plays a selective role in the retrieval of detailed contextual memories. Curr. Biol. 20 1336–1344. 10.1016/j.cub.2010.06.068 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Winocur G., Moscovitch M. (2011). Memory transformation and systems consolidation. J. Int. Neuropsychol. Soc. 17 766–780. 10.1017/S1355617711000683 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Wolf R. C., Vasic N., Walter H. (2006). Differential activation of ventrolateral prefrontal cortex during working memory retrieval. Neuropsychologia 44 2558–2563. 10.1016/j.neuropsychologia.2006.05.015 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Xu Y., Chun M. M. (2006). Dissociable neural mechanisms supporting visual short-term memory for objects. Nature 440 91–95. 10.1038/nature04262 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Xu Y., Zou P., Cohen A. E. (2017). Voltage imaging with genetically encoded indicators. Curr. Opin. Chem. Biol. 39 1–10. 10.1016/j.cbpa.2017.04.005 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Yang P., Wang M., Luo C., Ni X., Li L. (2022). Dissociable causal roles of the frontal and parietal cortices in the effect of object location on object identity detection: a TMS study. Exp. Brain Res. 240 1445–1457. 10.1007/s00221-022-06344-4 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Yu R., Han B., Wu X., Wei G., Zhang J., Ding M., et al. (2023). Dual-functional network regulation underlies the Central Executive System in working memory. Neuroscience 524 158–180. 10.1016/j.neuroscience.2023.05.025 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Yu S., Rempel S., Gholamipourbarogh N., Beste C. (2022). A ventral stream-prefrontal cortex processing cascade enables working memory gating dynamics. Commun. Biol. 5 : 1086 . 10.1038/s42003-022-04048-7 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Zeithamova D., Preston A. R. (2010). Flexible memories: differential roles for medial temporal lobe and prefrontal cortex in cross-episode binding. J. Neurosci. 30 14676–14684. 10.1523/jneurosci.3250-10.2010 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Zhu Y., Zeng Y., Ren J., Zhang L., Chen C., Fernandez G., et al. (2022). Emotional learning retroactively promotes memory integration through rapid neural reactivation and reorganization. eLife 11 : e60190 . 10.7554/elife.60190 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Zielinski M. C., Tang W., Jadhav S. P. (2018). The role of replay and Theta sequences in mediating hippocampal-prefrontal interactions for memory and cognition. Hippocampus 30 60–72. 10.1002/hipo.22821 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Zlotnik G., Vansintjan A. (2019). Memory: an extended definition. Front. Psychol. 10 : 2523 . 10.3389/fpsyg.2019.02523 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
Featured Topics
Featured series.
A series of random questions answered by Harvard experts.
Explore the Gazette
Read the latest.

‘Harvard Thinking’: Is AI friend or foe? Wrong question.

Getting ahead of dyslexia

Why AI fairness conversations must include disabled people
Making memories.
Kevin Jiang
HMS Communications
Study sheds light on how neurons form long-term memories
On a late summer day in 1953, a young man who would soon be known as patient H.M. underwent experimental surgery. In an attempt to treat his debilitating seizures, a surgeon removed portions of his brain, including part of a structure called the hippocampus. The seizures stopped.
Unfortunately, for patient H.M., so too did time. When he woke up after surgery, he could no longer form new long-term memories, despite retaining normal cognitive abilities, language and short-term working memory. Patient H.M.’s condition ultimately revealed that the brain’s ability to create long-term memories is a distinct process that depends on the hippocampus.
Scientists had discovered where memories are made. But how they are made remained unknown.
Now, neuroscientists at Harvard Medical School (HMS) have taken a decisive step in the quest to understand the biology of long-term memory and find ways to intervene when memory deficits occur with age or disease.
Reporting in Nature on Dec. 9, they describe a newly identified mechanism that neurons in the adult mouse hippocampus use to regulate signals they receive from other neurons, in a process that appears critical for memory consolidation and recall.
The study was led by Lynn Yap , HMS graduate student in neurobiology, and Michael Greenberg , chair of neurobiology in the Blavatnik Institute at HMS.
“If we can better understand this process, we will have new handles on memory and how to intervene when things go wrong, …” Michael Greenberg, Blavatnik Institute at HMS
“Memory is essential to all aspects of human existence. The question of how we encode memories that last a lifetime is a fundamental one, and our study gets to the very heart of this phenomenon,” said Greenberg, the HMS Nathan Marsh Pusey Professor of Neurobiology and study corresponding author.
The researchers observed that new experiences activate sparse populations of neurons in the hippocampus that express two genes, Fos and Scg2. These genes allow neurons to fine-tune inputs from so-called inhibitory interneurons, cells that dampen neuronal excitation. In this way, small groups of disparate neurons may form persistent networks with coordinated activity in response to an experience.
“This mechanism likely allows neurons to better talk to each other so that the next time a memory needs to be recalled, the neurons fire more synchronously,” Yap said. “We think coincident activation of this Fos-mediated circuit is potentially a necessary feature for memory consolidation, for example, during sleep, and also memory recall in the brain.”
Circuit orchestration
In order to form memories, the brain must somehow wire an experience into neurons so that when these neurons are reactivated, the initial experience can be recalled. In their study, Greenberg, Yap and team set out to explore this process by looking at the gene Fos.
First described in neuronal cells by Greenberg and colleagues in 1986, Fos is expressed within minutes after a neuron is activated. Scientists have taken advantage of this property, using Fos as a marker of recent neuronal activity to identify brain cells that regulate thirst, torpor , and many other behaviors.
Scientists hypothesized that Fos might play a critical role in learning and memory, but for decades, the precise function of the gene has remained a mystery.
To investigate, the researchers exposed mice to new environments and looked at pyramidal neurons, the principal cells of the hippocampus. They found that relatively sparse populations of neurons expressed Fos after exposure to a new experience. Next, they prevented these neurons from expressing Fos, using a virus-based tool delivered to a specific area of the hippocampus, which left other cells unaffected.
Mice that had Fos blocked in this manner showed significant memory deficits when assessed in a maze that required them to recall spatial details, indicating that the gene plays a critical role in memory formation.
The researchers studied the differences between neurons that expressed Fos and those that did not. Using optogenetics to turn inputs from different nearby neurons on or off, they discovered that the activity of Fos-expressing neurons was most strongly affected by two types of interneurons.
Neurons expressing Fos were found to receive increased activity-dampening, or inhibitory, signals from one distinct type of interneuron and decreased inhibitory signals from another type. These signaling patterns disappeared in neurons with blocked Fos expression.
“What’s critical about these interneurons is that they can regulate when and how much individual Fos-activated neurons fire, and also when they fire relative to other neurons in the circuit,” Yap said. “We think that at long last we have a handle on how Fos may in fact support memory processes, specifically by orchestrating this type of circuit plasticity in the hippocampus.”
Imagine the day
The researchers further probed the function of Fos, which codes for a transcription factor protein that regulates other genes. They used single-cell sequencing and additional genomic screens to identify genes activated by Fos and found that one gene in particular, Scg2, played a critical role in regulating inhibitory signals.
In mice with experimentally silenced Scg2, Fos-activated neurons in the hippocampus displayed a defect in signaling from both types of interneurons. These mice also had defects in theta and gamma rhythms, brain properties thought to be critical features of learning and memory.
Previous studies had shown that Scg2 codes for a neuropeptide protein that can be cleaved into four distinct forms, which are then secreted. In the current study, Yap and colleagues discovered that neurons appear to use these neuropeptides to fine-tune inputs they receive from interneurons.
Together, the team’s experiments suggest that after a new experience, a small group of neurons simultaneously express Fos, activating Scg2 and its derived neuropeptides, in order to establish a coordinated network with its activity regulated by interneurons.
“When neurons are activated in the hippocampus after a new experience, they aren’t necessarily linked together in any particular way in advance,” Greenberg said. “But interneurons have very broad axonal arbors, meaning they can connect with and signal to many cells at once. This may be how a sparse group of neurons can be linked together to ultimately encode a memory.”
The study findings represent a possible molecular- and circuit-level mechanism for long-term memory. They shed new light on the fundamental biology of memory formation and have broad implications for diseases of memory dysfunction.
The researchers note, however, that while the results are an important step in our understanding of the inner workings of memory, numerous unanswered questions about the newly identified mechanisms remain.
“We’re not quite at the answer yet, but we can now see many of the next steps that need to be taken,” Greenberg said. “If we can better understand this process, we will have new handles on memory and how to intervene when things go wrong, whether in age-related memory loss or neurodegenerative disorders such as Alzheimer’s disease.”
The findings also represent the culmination of decades of research, even as they open new avenues of study that will likely take decades more to explore, Greenberg added.
“I arrived at Harvard in 1986, just as my paper describing the discovery that neuronal activity can turn on genes was published,” he said. “Since that time, I’ve been imagining the day when we would figure out how genes like Fos might contribute to long-term memory.”
Additional authors include Noah Pettit, Christopher Davis, M. Aurel Nagy, David Harmin, Emily Golden, Onur Dagliyan, Cindy Lin, Stephanie Rudolph, Nikhil Sharma, Eric Griffith, and Christopher Harvey.
The study was supported by the National Institutes of Health (grants R01NS028829, R01NS115965, R01NS089521, T32NS007473 and F32NS112455), a Stuart H.Q. and Victoria Quan fellowship, a Harvard Department of Neurobiology graduate fellowship, an Aramont Fund.
Share this article
You might like.
In podcast, a lawyer, computer scientist, and statistician debate ethics of artificial intelligence

Harvard lab’s research suggests at-risk kids can be identified before they ever struggle in school

Tech offers promise to help yet too often perpetuates ableism, say researchers. It doesn’t have to be this way.
Harvard announces return to required testing
Leading researchers cite strong evidence that testing expands opportunity
Yes, it’s exciting. Just don’t look at the sun.
Lab, telescope specialist details Harvard eclipse-viewing party, offers safety tips
For all the other Willie Jacks
‘Reservation Dogs’ star Paulina Alexis offers behind-the-scenes glimpse of hit show, details value of Native representation
- Search Menu
- Browse content in Arts and Humanities
- Browse content in Archaeology
- Anglo-Saxon and Medieval Archaeology
- Archaeological Methodology and Techniques
- Archaeology by Region
- Archaeology of Religion
- Archaeology of Trade and Exchange
- Biblical Archaeology
- Contemporary and Public Archaeology
- Environmental Archaeology
- Historical Archaeology
- History and Theory of Archaeology
- Industrial Archaeology
- Landscape Archaeology
- Mortuary Archaeology
- Prehistoric Archaeology
- Underwater Archaeology
- Urban Archaeology
- Zooarchaeology
- Browse content in Architecture
- Architectural Structure and Design
- History of Architecture
- Residential and Domestic Buildings
- Theory of Architecture
- Browse content in Art
- Art Subjects and Themes
- History of Art
- Industrial and Commercial Art
- Theory of Art
- Biographical Studies
- Byzantine Studies
- Browse content in Classical Studies
- Classical History
- Classical Philosophy
- Classical Mythology
- Classical Literature
- Classical Reception
- Classical Art and Architecture
- Classical Oratory and Rhetoric
- Greek and Roman Papyrology
- Greek and Roman Epigraphy
- Greek and Roman Law
- Greek and Roman Archaeology
- Late Antiquity
- Religion in the Ancient World
- Digital Humanities
- Browse content in History
- Colonialism and Imperialism
- Diplomatic History
- Environmental History
- Genealogy, Heraldry, Names, and Honours
- Genocide and Ethnic Cleansing
- Historical Geography
- History by Period
- History of Emotions
- History of Agriculture
- History of Education
- History of Gender and Sexuality
- Industrial History
- Intellectual History
- International History
- Labour History
- Legal and Constitutional History
- Local and Family History
- Maritime History
- Military History
- National Liberation and Post-Colonialism
- Oral History
- Political History
- Public History
- Regional and National History
- Revolutions and Rebellions
- Slavery and Abolition of Slavery
- Social and Cultural History
- Theory, Methods, and Historiography
- Urban History
- World History
- Browse content in Language Teaching and Learning
- Language Learning (Specific Skills)
- Language Teaching Theory and Methods
- Browse content in Linguistics
- Applied Linguistics
- Cognitive Linguistics
- Computational Linguistics
- Forensic Linguistics
- Grammar, Syntax and Morphology
- Historical and Diachronic Linguistics
- History of English
- Language Evolution
- Language Reference
- Language Acquisition
- Language Variation
- Language Families
- Lexicography
- Linguistic Anthropology
- Linguistic Theories
- Linguistic Typology
- Phonetics and Phonology
- Psycholinguistics
- Sociolinguistics
- Translation and Interpretation
- Writing Systems
- Browse content in Literature
- Bibliography
- Children's Literature Studies
- Literary Studies (Romanticism)
- Literary Studies (American)
- Literary Studies (Asian)
- Literary Studies (European)
- Literary Studies (Eco-criticism)
- Literary Studies (Modernism)
- Literary Studies - World
- Literary Studies (1500 to 1800)
- Literary Studies (19th Century)
- Literary Studies (20th Century onwards)
- Literary Studies (African American Literature)
- Literary Studies (British and Irish)
- Literary Studies (Early and Medieval)
- Literary Studies (Fiction, Novelists, and Prose Writers)
- Literary Studies (Gender Studies)
- Literary Studies (Graphic Novels)
- Literary Studies (History of the Book)
- Literary Studies (Plays and Playwrights)
- Literary Studies (Poetry and Poets)
- Literary Studies (Postcolonial Literature)
- Literary Studies (Queer Studies)
- Literary Studies (Science Fiction)
- Literary Studies (Travel Literature)
- Literary Studies (War Literature)
- Literary Studies (Women's Writing)
- Literary Theory and Cultural Studies
- Mythology and Folklore
- Shakespeare Studies and Criticism
- Browse content in Media Studies
- Browse content in Music
- Applied Music
- Dance and Music
- Ethics in Music
- Ethnomusicology
- Gender and Sexuality in Music
- Medicine and Music
- Music Cultures
- Music and Media
- Music and Religion
- Music and Culture
- Music Education and Pedagogy
- Music Theory and Analysis
- Musical Scores, Lyrics, and Libretti
- Musical Structures, Styles, and Techniques
- Musicology and Music History
- Performance Practice and Studies
- Race and Ethnicity in Music
- Sound Studies
- Browse content in Performing Arts
- Browse content in Philosophy
- Aesthetics and Philosophy of Art
- Epistemology
- Feminist Philosophy
- History of Western Philosophy
- Metaphysics
- Moral Philosophy
- Non-Western Philosophy
- Philosophy of Language
- Philosophy of Mind
- Philosophy of Perception
- Philosophy of Science
- Philosophy of Action
- Philosophy of Law
- Philosophy of Religion
- Philosophy of Mathematics and Logic
- Practical Ethics
- Social and Political Philosophy
- Browse content in Religion
- Biblical Studies
- Christianity
- East Asian Religions
- History of Religion
- Judaism and Jewish Studies
- Qumran Studies
- Religion and Education
- Religion and Health
- Religion and Politics
- Religion and Science
- Religion and Law
- Religion and Art, Literature, and Music
- Religious Studies
- Browse content in Society and Culture
- Cookery, Food, and Drink
- Cultural Studies
- Customs and Traditions
- Ethical Issues and Debates
- Hobbies, Games, Arts and Crafts
- Lifestyle, Home, and Garden
- Natural world, Country Life, and Pets
- Popular Beliefs and Controversial Knowledge
- Sports and Outdoor Recreation
- Technology and Society
- Travel and Holiday
- Visual Culture
- Browse content in Law
- Arbitration
- Browse content in Company and Commercial Law
- Commercial Law
- Company Law
- Browse content in Comparative Law
- Systems of Law
- Competition Law
- Browse content in Constitutional and Administrative Law
- Government Powers
- Judicial Review
- Local Government Law
- Military and Defence Law
- Parliamentary and Legislative Practice
- Construction Law
- Contract Law
- Browse content in Criminal Law
- Criminal Procedure
- Criminal Evidence Law
- Sentencing and Punishment
- Employment and Labour Law
- Environment and Energy Law
- Browse content in Financial Law
- Banking Law
- Insolvency Law
- History of Law
- Human Rights and Immigration
- Intellectual Property Law
- Browse content in International Law
- Private International Law and Conflict of Laws
- Public International Law
- IT and Communications Law
- Jurisprudence and Philosophy of Law
- Law and Politics
- Law and Society
- Browse content in Legal System and Practice
- Courts and Procedure
- Legal Skills and Practice
- Primary Sources of Law
- Regulation of Legal Profession
- Medical and Healthcare Law
- Browse content in Policing
- Criminal Investigation and Detection
- Police and Security Services
- Police Procedure and Law
- Police Regional Planning
- Browse content in Property Law
- Personal Property Law
- Study and Revision
- Terrorism and National Security Law
- Browse content in Trusts Law
- Wills and Probate or Succession
- Browse content in Medicine and Health
- Browse content in Allied Health Professions
- Arts Therapies
- Clinical Science
- Dietetics and Nutrition
- Occupational Therapy
- Operating Department Practice
- Physiotherapy
- Radiography
- Speech and Language Therapy
- Browse content in Anaesthetics
- General Anaesthesia
- Neuroanaesthesia
- Clinical Neuroscience
- Browse content in Clinical Medicine
- Acute Medicine
- Cardiovascular Medicine
- Clinical Genetics
- Clinical Pharmacology and Therapeutics
- Dermatology
- Endocrinology and Diabetes
- Gastroenterology
- Genito-urinary Medicine
- Geriatric Medicine
- Infectious Diseases
- Medical Toxicology
- Medical Oncology
- Pain Medicine
- Palliative Medicine
- Rehabilitation Medicine
- Respiratory Medicine and Pulmonology
- Rheumatology
- Sleep Medicine
- Sports and Exercise Medicine
- Community Medical Services
- Critical Care
- Emergency Medicine
- Forensic Medicine
- Haematology
- History of Medicine
- Browse content in Medical Skills
- Clinical Skills
- Communication Skills
- Nursing Skills
- Surgical Skills
- Browse content in Medical Dentistry
- Oral and Maxillofacial Surgery
- Paediatric Dentistry
- Restorative Dentistry and Orthodontics
- Surgical Dentistry
- Medical Ethics
- Medical Statistics and Methodology
- Browse content in Neurology
- Clinical Neurophysiology
- Neuropathology
- Nursing Studies
- Browse content in Obstetrics and Gynaecology
- Gynaecology
- Occupational Medicine
- Ophthalmology
- Otolaryngology (ENT)
- Browse content in Paediatrics
- Neonatology
- Browse content in Pathology
- Chemical Pathology
- Clinical Cytogenetics and Molecular Genetics
- Histopathology
- Medical Microbiology and Virology
- Patient Education and Information
- Browse content in Pharmacology
- Psychopharmacology
- Browse content in Popular Health
- Caring for Others
- Complementary and Alternative Medicine
- Self-help and Personal Development
- Browse content in Preclinical Medicine
- Cell Biology
- Molecular Biology and Genetics
- Reproduction, Growth and Development
- Primary Care
- Professional Development in Medicine
- Browse content in Psychiatry
- Addiction Medicine
- Child and Adolescent Psychiatry
- Forensic Psychiatry
- Learning Disabilities
- Old Age Psychiatry
- Psychotherapy
- Browse content in Public Health and Epidemiology
- Epidemiology
- Public Health
- Browse content in Radiology
- Clinical Radiology
- Interventional Radiology
- Nuclear Medicine
- Radiation Oncology
- Reproductive Medicine
- Browse content in Surgery
- Cardiothoracic Surgery
- Gastro-intestinal and Colorectal Surgery
- General Surgery
- Neurosurgery
- Paediatric Surgery
- Peri-operative Care
- Plastic and Reconstructive Surgery
- Surgical Oncology
- Transplant Surgery
- Trauma and Orthopaedic Surgery
- Vascular Surgery
- Browse content in Science and Mathematics
- Browse content in Biological Sciences
- Aquatic Biology
- Biochemistry
- Bioinformatics and Computational Biology
- Developmental Biology
- Ecology and Conservation
- Evolutionary Biology
- Genetics and Genomics
- Microbiology
- Molecular and Cell Biology
- Natural History
- Plant Sciences and Forestry
- Research Methods in Life Sciences
- Structural Biology
- Systems Biology
- Zoology and Animal Sciences
- Browse content in Chemistry
- Analytical Chemistry
- Computational Chemistry
- Crystallography
- Environmental Chemistry
- Industrial Chemistry
- Inorganic Chemistry
- Materials Chemistry
- Medicinal Chemistry
- Mineralogy and Gems
- Organic Chemistry
- Physical Chemistry
- Polymer Chemistry
- Study and Communication Skills in Chemistry
- Theoretical Chemistry
- Browse content in Computer Science
- Artificial Intelligence
- Computer Architecture and Logic Design
- Game Studies
- Human-Computer Interaction
- Mathematical Theory of Computation
- Programming Languages
- Software Engineering
- Systems Analysis and Design
- Virtual Reality
- Browse content in Computing
- Business Applications
- Computer Security
- Computer Games
- Computer Networking and Communications
- Digital Lifestyle
- Graphical and Digital Media Applications
- Operating Systems
- Browse content in Earth Sciences and Geography
- Atmospheric Sciences
- Environmental Geography
- Geology and the Lithosphere
- Maps and Map-making
- Meteorology and Climatology
- Oceanography and Hydrology
- Palaeontology
- Physical Geography and Topography
- Regional Geography
- Soil Science
- Urban Geography
- Browse content in Engineering and Technology
- Agriculture and Farming
- Biological Engineering
- Civil Engineering, Surveying, and Building
- Electronics and Communications Engineering
- Energy Technology
- Engineering (General)
- Environmental Science, Engineering, and Technology
- History of Engineering and Technology
- Mechanical Engineering and Materials
- Technology of Industrial Chemistry
- Transport Technology and Trades
- Browse content in Environmental Science
- Applied Ecology (Environmental Science)
- Conservation of the Environment (Environmental Science)
- Environmental Sustainability
- Environmentalist Thought and Ideology (Environmental Science)
- Management of Land and Natural Resources (Environmental Science)
- Natural Disasters (Environmental Science)
- Nuclear Issues (Environmental Science)
- Pollution and Threats to the Environment (Environmental Science)
- Social Impact of Environmental Issues (Environmental Science)
- History of Science and Technology
- Browse content in Materials Science
- Ceramics and Glasses
- Composite Materials
- Metals, Alloying, and Corrosion
- Nanotechnology
- Browse content in Mathematics
- Applied Mathematics
- Biomathematics and Statistics
- History of Mathematics
- Mathematical Education
- Mathematical Finance
- Mathematical Analysis
- Numerical and Computational Mathematics
- Probability and Statistics
- Pure Mathematics
- Browse content in Neuroscience
- Cognition and Behavioural Neuroscience
- Development of the Nervous System
- Disorders of the Nervous System
- History of Neuroscience
- Invertebrate Neurobiology
- Molecular and Cellular Systems
- Neuroendocrinology and Autonomic Nervous System
- Neuroscientific Techniques
- Sensory and Motor Systems
- Browse content in Physics
- Astronomy and Astrophysics
- Atomic, Molecular, and Optical Physics
- Biological and Medical Physics
- Classical Mechanics
- Computational Physics
- Condensed Matter Physics
- Electromagnetism, Optics, and Acoustics
- History of Physics
- Mathematical and Statistical Physics
- Measurement Science
- Nuclear Physics
- Particles and Fields
- Plasma Physics
- Quantum Physics
- Relativity and Gravitation
- Semiconductor and Mesoscopic Physics
- Browse content in Psychology
- Affective Sciences
- Clinical Psychology
- Cognitive Psychology
- Cognitive Neuroscience
- Criminal and Forensic Psychology
- Developmental Psychology
- Educational Psychology
- Evolutionary Psychology
- Health Psychology
- History and Systems in Psychology
- Music Psychology
- Neuropsychology
- Organizational Psychology
- Psychological Assessment and Testing
- Psychology of Human-Technology Interaction
- Psychology Professional Development and Training
- Research Methods in Psychology
- Social Psychology
- Browse content in Social Sciences
- Browse content in Anthropology
- Anthropology of Religion
- Human Evolution
- Medical Anthropology
- Physical Anthropology
- Regional Anthropology
- Social and Cultural Anthropology
- Theory and Practice of Anthropology
- Browse content in Business and Management
- Business Ethics
- Business Strategy
- Business History
- Business and Technology
- Business and Government
- Business and the Environment
- Comparative Management
- Corporate Governance
- Corporate Social Responsibility
- Entrepreneurship
- Health Management
- Human Resource Management
- Industrial and Employment Relations
- Industry Studies
- Information and Communication Technologies
- International Business
- Knowledge Management
- Management and Management Techniques
- Operations Management
- Organizational Theory and Behaviour
- Pensions and Pension Management
- Public and Nonprofit Management
- Strategic Management
- Supply Chain Management
- Browse content in Criminology and Criminal Justice
- Criminal Justice
- Criminology
- Forms of Crime
- International and Comparative Criminology
- Youth Violence and Juvenile Justice
- Development Studies
- Browse content in Economics
- Agricultural, Environmental, and Natural Resource Economics
- Asian Economics
- Behavioural Finance
- Behavioural Economics and Neuroeconomics
- Econometrics and Mathematical Economics
- Economic History
- Economic Systems
- Economic Methodology
- Economic Development and Growth
- Financial Markets
- Financial Institutions and Services
- General Economics and Teaching
- Health, Education, and Welfare
- History of Economic Thought
- International Economics
- Labour and Demographic Economics
- Law and Economics
- Macroeconomics and Monetary Economics
- Microeconomics
- Public Economics
- Urban, Rural, and Regional Economics
- Welfare Economics
- Browse content in Education
- Adult Education and Continuous Learning
- Care and Counselling of Students
- Early Childhood and Elementary Education
- Educational Equipment and Technology
- Educational Strategies and Policy
- Higher and Further Education
- Organization and Management of Education
- Philosophy and Theory of Education
- Schools Studies
- Secondary Education
- Teaching of a Specific Subject
- Teaching of Specific Groups and Special Educational Needs
- Teaching Skills and Techniques
- Browse content in Environment
- Applied Ecology (Social Science)
- Climate Change
- Conservation of the Environment (Social Science)
- Environmentalist Thought and Ideology (Social Science)
- Natural Disasters (Environment)
- Social Impact of Environmental Issues (Social Science)
- Browse content in Human Geography
- Cultural Geography
- Economic Geography
- Political Geography
- Browse content in Interdisciplinary Studies
- Communication Studies
- Museums, Libraries, and Information Sciences
- Browse content in Politics
- African Politics
- Asian Politics
- Chinese Politics
- Comparative Politics
- Conflict Politics
- Elections and Electoral Studies
- Environmental Politics
- European Union
- Foreign Policy
- Gender and Politics
- Human Rights and Politics
- Indian Politics
- International Relations
- International Organization (Politics)
- International Political Economy
- Irish Politics
- Latin American Politics
- Middle Eastern Politics
- Political Behaviour
- Political Economy
- Political Institutions
- Political Methodology
- Political Communication
- Political Philosophy
- Political Sociology
- Political Theory
- Politics and Law
- Public Policy
- Public Administration
- Quantitative Political Methodology
- Regional Political Studies
- Russian Politics
- Security Studies
- State and Local Government
- UK Politics
- US Politics
- Browse content in Regional and Area Studies
- African Studies
- Asian Studies
- East Asian Studies
- Japanese Studies
- Latin American Studies
- Middle Eastern Studies
- Native American Studies
- Scottish Studies
- Browse content in Research and Information
- Research Methods
- Browse content in Social Work
- Addictions and Substance Misuse
- Adoption and Fostering
- Care of the Elderly
- Child and Adolescent Social Work
- Couple and Family Social Work
- Developmental and Physical Disabilities Social Work
- Direct Practice and Clinical Social Work
- Emergency Services
- Human Behaviour and the Social Environment
- International and Global Issues in Social Work
- Mental and Behavioural Health
- Social Justice and Human Rights
- Social Policy and Advocacy
- Social Work and Crime and Justice
- Social Work Macro Practice
- Social Work Practice Settings
- Social Work Research and Evidence-based Practice
- Welfare and Benefit Systems
- Browse content in Sociology
- Childhood Studies
- Community Development
- Comparative and Historical Sociology
- Economic Sociology
- Gender and Sexuality
- Gerontology and Ageing
- Health, Illness, and Medicine
- Marriage and the Family
- Migration Studies
- Occupations, Professions, and Work
- Organizations
- Population and Demography
- Race and Ethnicity
- Social Theory
- Social Movements and Social Change
- Social Research and Statistics
- Social Stratification, Inequality, and Mobility
- Sociology of Religion
- Sociology of Education
- Sport and Leisure
- Urban and Rural Studies
- Browse content in Warfare and Defence
- Defence Strategy, Planning, and Research
- Land Forces and Warfare
- Military Administration
- Military Life and Institutions
- Naval Forces and Warfare
- Other Warfare and Defence Issues
- Peace Studies and Conflict Resolution
- Weapons and Equipment

The Oxford Handbook of Memory
- Cite Icon Cite
- Permissions Icon Permissions
Due to the advent of neuropsychology, it has become clear that there is a multiplicity of memory systems or, at the very least, of dissociably different modes of processing memory in the brain. As the Oxford Handbook of Memory demonstrates, the frontier of memory research has been enriched by breakthroughs of the last decades, with lines of continuity and important departures, and it will continue to be enriched by changes in technology that will propel future research. In turn, such changes are beginning to impact the legal and professional therapeutic professions and will have considerable future significance in realms outside of psychology and memory research. Endel Tulving and Fergus Craik, two world-class experts on memory, provide this handbook as a roadmap to the huge and unwieldy field of memory research. By enlisting an eminent group of researchers, they are able to offer insight into breakthroughs for the work that lies ahead. The outline is comprehensive and covers such topics as the development of memory, the contents of memory, memory in the laboratory and in everyday use, memory in decline, the organization of memory, and theories of memory.
Signed in as
Institutional accounts.
- Google Scholar Indexing
- GoogleCrawler [DO NOT DELETE]
Personal account
- Sign in with email/username & password
- Get email alerts
- Save searches
- Purchase content
- Activate your purchase/trial code
Institutional access
- Sign in with a library card Sign in with username/password Recommend to your librarian
- Institutional account management
- Get help with access
Access to content on Oxford Academic is often provided through institutional subscriptions and purchases. If you are a member of an institution with an active account, you may be able to access content in one of the following ways:
IP based access
Typically, access is provided across an institutional network to a range of IP addresses. This authentication occurs automatically, and it is not possible to sign out of an IP authenticated account.
Sign in through your institution
Choose this option to get remote access when outside your institution. Shibboleth/Open Athens technology is used to provide single sign-on between your institution’s website and Oxford Academic.
- Click Sign in through your institution.
- Select your institution from the list provided, which will take you to your institution's website to sign in.
- When on the institution site, please use the credentials provided by your institution. Do not use an Oxford Academic personal account.
- Following successful sign in, you will be returned to Oxford Academic.
If your institution is not listed or you cannot sign in to your institution’s website, please contact your librarian or administrator.
Sign in with a library card
Enter your library card number to sign in. If you cannot sign in, please contact your librarian.
Society Members
Society member access to a journal is achieved in one of the following ways:
Sign in through society site
Many societies offer single sign-on between the society website and Oxford Academic. If you see ‘Sign in through society site’ in the sign in pane within a journal:
- Click Sign in through society site.
- When on the society site, please use the credentials provided by that society. Do not use an Oxford Academic personal account.
If you do not have a society account or have forgotten your username or password, please contact your society.
Sign in using a personal account
Some societies use Oxford Academic personal accounts to provide access to their members. See below.
A personal account can be used to get email alerts, save searches, purchase content, and activate subscriptions.
Some societies use Oxford Academic personal accounts to provide access to their members.
Viewing your signed in accounts
Click the account icon in the top right to:
- View your signed in personal account and access account management features.
- View the institutional accounts that are providing access.
Signed in but can't access content
Oxford Academic is home to a wide variety of products. The institutional subscription may not cover the content that you are trying to access. If you believe you should have access to that content, please contact your librarian.
For librarians and administrators, your personal account also provides access to institutional account management. Here you will find options to view and activate subscriptions, manage institutional settings and access options, access usage statistics, and more.
Our books are available by subscription or purchase to libraries and institutions.
- About Oxford Academic
- Publish journals with us
- University press partners
- What we publish
- New features
- Open access
- Rights and permissions
- Accessibility
- Advertising
- Media enquiries
- Oxford University Press
- Oxford Languages
- University of Oxford
Oxford University Press is a department of the University of Oxford. It furthers the University's objective of excellence in research, scholarship, and education by publishing worldwide
- Copyright © 2024 Oxford University Press
- Cookie settings
- Cookie policy
- Privacy policy
- Legal notice
This Feature Is Available To Subscribers Only
Sign In or Create an Account
This PDF is available to Subscribers Only
For full access to this pdf, sign in to an existing account, or purchase an annual subscription.
Thank you for visiting nature.com. You are using a browser version with limited support for CSS. To obtain the best experience, we recommend you use a more up to date browser (or turn off compatibility mode in Internet Explorer). In the meantime, to ensure continued support, we are displaying the site without styles and JavaScript.
- View all journals
- Explore content
- About the journal
- Publish with us
- Sign up for alerts
- Perspective
- Published: 22 December 2023
Optimizing the methodology of human sleep and memory research
- Dezső Németh ORCID: orcid.org/0000-0002-9629-5856 1 , 2 , 3 na1 ,
- Emilie Gerbier 4 na1 ,
- Jan Born 5 ,
- Timothy Rickard 6 ,
- Susanne Diekelmann 5 , 7 ,
- Stuart Fogel ORCID: orcid.org/0000-0002-3227-5370 8 ,
- Lisa Genzel 9 ,
- Alexander Prehn-Kristensen 10 , 11 ,
- Jessica Payne 12 ,
- Martin Dresler ORCID: orcid.org/0000-0001-7441-3818 13 ,
- Peter Simor 2 , 14 ,
- Stephanie Mazza 15 ,
- Kerstin Hoedlmoser 16 ,
- Perrine Ruby 1 ,
- Rebecca M. C. Spencer 17 ,
- Genevieve Albouy 18 ,
- Teodóra Vékony 1 ,
- Manuel Schabus ORCID: orcid.org/0000-0001-5899-8772 16 &
- Karolina Janacsek 2 , 19 na1
Nature Reviews Psychology volume 3 , pages 123–137 ( 2024 ) Cite this article
1030 Accesses
15 Altmetric
Metrics details
- Human behaviour
- Learning and memory
- Long-term memory
Understanding the complex relationship between sleep and memory consolidation is a major challenge in cognitive neuroscience and psychology. Many studies suggest that sleep triggers off-line memory processes, resulting in less forgetting of declarative memory and performance stabilization in non-declarative memory. However, the role of sleep in human memory consolidation is still under considerable debate, and numerous contradictory and non-replicable findings have been reported. Methodological issues related to experimental designs, task characteristics and measurements, and data-analysis practices all influence the effects that are observed and their interpretation. In this Perspective, we review methodological issues in sleep and memory studies and suggest constructive solutions to address them. We believe that implementing these solutions in future sleep and memory research will substantially advance the field and improve understanding of the specific role of sleep in memory consolidation.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 digital issues and online access to articles
55,14 € per year
only 4,60 € per issue
Rent or buy this article
Prices vary by article type
Prices may be subject to local taxes which are calculated during checkout
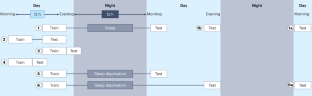
Similar content being viewed by others

The relationship between subjective sleep quality and cognitive performance in healthy young adults: Evidence from three empirical studies
Zsófia Zavecz, Tamás Nagy, … Karolina Janacsek

Individual sleep need is flexible and dynamically related to cognitive function
Anders M. Fjell & Kristine B. Walhovd

Offline memory consolidation during waking rest
Erin J. Wamsley
Palagini, L. & Rosenlicht, N. Sleep, dreaming, and mental health: a review of historical and neurobiological perspectives. Sleep. Med. Rev. 15 , 179–186 (2011).
Article PubMed Google Scholar
Simor, P., Sifuentes-Ortega, R. & Peigneux, P. in Sleep Medicine Textbook (eds Bassetti, C., McNicholas, W., Paunio, T. & Peigneux, P.) Ch. A.5 (European Sleep Research Society, 2021).
Walker, M. P. The role of sleep in cognition and emotion. Ann. NY Acad. Sci. 1156 , 168–197 (2009).
Article PubMed ADS Google Scholar
Shochat, T. Impact of lifestyle and technology developments on sleep. Nat. Sci. Sleep 4 , 19–31 (2012).
Article PubMed PubMed Central Google Scholar
Bixler, E. Sleep and society: an epidemiological perspective. Sleep. Med. 10 , S3 (2009).
Matricciani, L. et al. Past, present, and future: trends in sleep duration and implications for public health. Sleep. Health 3 , 317–323 (2017).
Hale, L. & Guan, S. Screen time and sleep among school-aged children and adolescents: a systematic literature review. Sleep. Med. Rev. 21 , 50–58 (2015).
Ackermann, S. & Rasch, B. Differential effects of non-REM and REM sleep on memory consolidation? Curr. Neurol. Neurosci. Rep. 14 , 430 (2014).
Antony, J. W., Schönauer, M., Staresina, B. P. & Cairney, S. A. Sleep spindles and memory reprocessing. Trends Neurosci. 42 , 1–3 (2019).
Article CAS PubMed Google Scholar
Boyce, R., Williams, S. & Adamantidis, A. REM sleep and memory. Curr. Opin. Neurobiol. 44 , 167–177 (2017).
Diekelmann, S. & Born, J. The memory function of sleep. Nat. Rev. Neurosci. 11 , 114–126 (2010).
Diekelmann, S., Wilhelm, I. & Born, J. The whats and whens of sleep-dependent memory consolidation. Sleep. Med. Rev. 13 , 309–321 (2009).
Feld, G. B. & Born, J. Sculpting memory during sleep: concurrent consolidation and forgetting. Curr. Opin. Neurobiol. 44 , 20–27 (2017).
Lewis, P. A. & Durrant, S. J. Overlapping memory replay during sleep builds cognitive schemata. Trends Cogn. Sci. 15 , 343–351 (2011).
Mednick, S. C., Cai, D. J., Shuman, T., Anagnostaras, S. & Wixted, J. T. An opportunistic theory of cellular and systems consolidation. Trends Neurosci. 34 , 504–514 (2011).
Article CAS PubMed PubMed Central Google Scholar
Saletin, J. M. & Walker, M. P. Nocturnal mnemonics: sleep and hippocampal memory processing. Front. Neurol. 3 , 57 (2012).
Article Google Scholar
Siegel, D. J. Memory: an overview, with emphasis on developmental, interpersonal, and neurobiological aspects. J. Am. Acad. Child. Adolesc. Psychiat. 40 , 997–1011 (2001).
Article CAS Google Scholar
Stickgold, R. & Walker, M. P. Memory consolidation and reconsolidation: what is the role of sleep? Trends Neurosci. 28 , 408–415 (2005).
Stickgold, R. & Walker, M. P. Sleep-dependent memory triage: evolving generalization through selective processing. Nat. Neurosci. 16 , 139–145 (2013).
Tononi, G. & Cirelli, C. Sleep function and synaptic homeostasis. Sleep. Med. Rev. 10 , 49–62 (2006).
Tononi, G. & Cirelli, C. Sleep and the price of plasticity: from synaptic and cellular homeostasis to memory consolidation and integration. Neuron 81 , 12–34 (2014).
Walker, M. P. A refined model of sleep and the time course of memory formation. Behav. Brain Sci. 28 , 51–64 (2005).
Schönauer, M., Grätsch, M. & Gais, S. Evidence for two distinct sleep-related long-term memory consolidation processes. Cortex 63 , 68–78 (2015).
Plihal, W. & Born, J. Effects of early and late nocturnal sleep on declarative and procedural memory. J. Cogn. Neurosci. 9 , 534–547 (1997).
Feld, G. B., Weis, P. P. & Born, J. The limited capacity of sleep-dependent memory consolidation. Front. Psychol. 7 , 204960 (2016).
Diekelmann, S., Born, J. & Wagner, U. Sleep enhances false memories depending on general memory performance. Behav. Brain Res. 208 , 425–429 (2010).
Abel, M. & Bäuml, K. H. T. Retrieval-induced forgetting, delay, and sleep. Memory 20 , 420–428 (2012).
Payne, J. D. et al. Napping and the selective consolidation of negative aspects of scenes. Emotion 15 , 176–186 (2015).
Cairney, S. A., Durrant, S. J., Power, R. & Lewis, P. A. Complementary roles of slow-wave sleep and rapid eye movement sleep in emotional memory consolidation. Cereb. Cortex 25 , 1565–1575 (2015).
Rasch, B., Büchel, C., Gais, S. & Born, J. Odor cues during slow-wave sleep prompt declarative memory consolidation. Science 315 , 1426–1429 (2007).
Article CAS PubMed ADS Google Scholar
Rudoy, J. D., Voss, J. L., Westerberg, C. E. & Paller, K. A. Strengthening individual memories by reactivating them during sleep. Science 326 , 1079 (2009).
Article CAS PubMed PubMed Central ADS Google Scholar
Gais, S., Lucas, B. & Born, J. Sleep after learning aids memory recall. Learn. Mem. 13 , 259–262 (2006).
Walker, M. P., Brakefield, T., Hobson, J. A. & Stickgold, R. Dissociable stages of human memory consolidation and reconsolidation. Nature 425 , 616–620 (2003).
King, B. R., Hoedlmoser, K., Hirschauer, F., Dolfen, N. & Albouy, G. Sleeping on the motor engram: the multifaceted nature of sleep-related motor memory consolidation. Neurosci. Biobehav. Rev. 80 , 1–22 (2017).
Rasch, B. & Born, J. About sleep’s role in memory. Physiol. Rev. 93 , 681–766 (2013).
Fischer, S., Hallschmid, M., Elsner, A. L. & Born, J. Sleep forms memory for finger skills. Proc. Natl Acad. Sci. USA 99 , 11987–11991 (2002).
Born, J. & Wilhelm, I. System consolidation of memory during sleep. Psychol. Res. 76 , 192–203 (2012).
Vorster, A. P. & Born, J. Sleep and memory in mammals, birds and invertebrates. Neurosci. Biobehav. Rev. 50 , 103–119 (2015).
Pan, S. C. & Rickard, T. C. Sleep and motor learning: is there room for consolidation? Psychol. Bull. 141 , 812–834 (2015).
Mantua, J. Sleep physiology correlations and human memory consolidation: where do we go from here? Sleep 41 , zsx204 (2018).
Vertes, R. P. & Siegel, J. M. Time for the sleep community to take a critical look at the purported role of sleep in memory processing. Sleep 28 , 1228–1229 (2005).
Dastgheib, M., Kulanayagam, A. & Dringenberg, H. C. Is the role of sleep in memory consolidation overrated? Neurosci. Biobehav. Rev. 140 , 104799 (2022).
Cordi, M. J. & Rasch, B. How robust are sleep-mediated memory benefits? Curr. Opin. Neurobiol. 67 , 1–7 (2021).
Voderholzer, U. et al. Sleep restriction over several days does not affect long-term recall of declarative and procedural memories in adolescents. Sleep Med. 12 , 170–178 (2011).
Payne, J. D. et al. The role of sleep in false memory formation. Neurobiol. Learn. Mem. 92 , 327–334 (2009).
Ujma, P. P. Meta-analytic evidence suggests no correlation between sleep spindles and memory. Preprint at PsyArXiv https://doi.org/10.31234/osf.io/hjdex (2022).
Krakauer, J. W., Ghazanfar, A. A., Gomez-Marin, A., MacIver, M. A. & Poeppel, D. Neuroscience needs behavior: correcting a reductionist bias. Neuron 93 , 480–490 (2017).
Niv, Y. The primacy of behavioral research for understanding the brain. Behav. Neurosci. 135 , 601–609 (2021).
Peigneux, P., Fogel, S. & Smith, C. Memory processing in relation to sleep. Princip. Pract. Sleep Med . https://doi.org/10.1016/b978-0-323-24288-2.00022-2 (2017).
Schmidt, C., Collette, F., Cajochen, C. & Peigneux, P. A time to think: circadian rhythms in human cognition. Cogn. Neuropsychol. 24 , 755–789 (2007).
Barner, C., Schmid, S. R. & Diekelmann, S. Time-of-day effects on prospective memory. Behav. Brain Res. 376 , 112179 (2019).
Scullin, M. K. & McDaniel, M. A. Remembering to execute a goal: sleep on it! Psychol. Sci. 21 , 1028–1035 (2010).
Keisler, A., Ashe, J. & Willingham, D. T. Time of day accounts for overnight improvement in sequence learning. Learn. Mem. 14 , 669–672 (2007).
Taillard, J., Gronfier, C., Bioulac, S., Philip, P. & Sagaspe, P. Sleep in normal aging, homeostatic and circadian regulation and vulnerability to sleep deprivation. Brain Sci. 11 , 1003 (2021).
Tucker, M. A. et al. The relative impact of sleep and circadian drive on motor skill acquisition and memory consolidation. Sleep 40 , zsx096 (2017).
Payne, J. D., Ellenbogen, J. M., Walker, M. P. & Stickgold, R. in Learning and Memory: Comprehensive Reference 1st edn, Vol. 2 (eds Byrne, J. H.) 663–685 (Academic, 2008).
Dresler, M. et al. Neuroscience-driven discovery and development of sleep therapeutics. Pharmacol. Ther. 141 , 300–334 (2014).
Fenn, K. M., Nusbaum, H. C. & Margoliash, D. Consolidation during sleep of perceptual learning of spoken language. Nature 425 , 614–616 (2003).
Hallgató, E., Gyori-Dani, D., Pekár, J., Janacsek, K. & Nemeth, D. The differential consolidation of perceptual and motor learning in skill acquisition. Cortex 49 , 1073–1081 (2013).
Talamini, L. M., Nieuwenhuis, I. L. C., Takashima, A. & Jensen, O. Sleep directly following learning benefits consolidation of spatial associative memory. Learn. Mem. 15 , 233–237 (2008).
Tucker, M., McKinley, S. & Stickgold, R. Sleep optimizes motor skill in older adults. J. Am. Geriatr. Soc. 59 , 603–609 (2011).
Gais, S., Rasch, B., Dahmen, J. C., Sara, S. & Born, J. The memory function of noradrenergic activity in non-REM sleep. J. Cogn. Neurosci. 23 , 2582–2592 (2011).
Csabi, E., Varszegi-Schulz, M., Janacsek, K., Malecek, N. & Nemeth, D. The consolidation of implicit sequence memory in obstructive sleep apnea. PLoS One 9 , e109010 (2014).
Article PubMed PubMed Central ADS Google Scholar
Backhaus, J. et al. Impaired declarative memory consolidation during sleep in patients with primary insomnia: influence of sleep architecture and nocturnal cortisol release. Biol. Psychiat. 60 , 1324–1330 (2006).
Ahuja, S. et al. Role of normal sleep and sleep apnea in human memory processing. Nat. Sci. Sleep. 10 , 255–269 (2018).
Cellini, N. Memory consolidation in sleep disorders. Sleep. Med. Rev. 35 , 101–112 (2017).
Wallace, A. & Bucks, R. S. Memory and obstructive sleep apnea: a meta-analysis. Sleep 36 , 203–220 (2013).
PubMed PubMed Central Google Scholar
Csábi, E., Benedek, P., Janacsek, K., Katona, G. & Nemeth, D. Sleep disorder in childhood impairs declarative but not nondeclarative forms of learning. J. Clin. Exp. Neuropsychol. 35 , 677–685 (2013).
Rosenzweig, I. et al. Sleep apnoea and the brain: a complex relationship. Lancet Respir. Med. 3 , 404–414 (2015).
Wulff, K., Gatti, S., Wettstein, J. G. & Foster, R. G. Sleep and circadian rhythm disruption in psychiatric and neurodegenerative disease. Nat. Rev. Neurosci. 11 , 589–599 (2010).
Li, J., Vitiello, M. V. & Gooneratne, N. S. Sleep in normal aging. Sleep. Med. Clin. 13 , 1–11 (2018).
Bennett, I. J., Howard, J. H. & Howard, D. V. Age-related differences in implicit learning of subtle third-order sequential structure. J. Gerontol. Ser. B 62 , P98–P103 (2007).
Nissen, C. et al. Sleep-related memory consolidation in primary insomnia. J. Sleep. Res. 20 , 129–136 (2011).
Ellenbogen, J. M., Payne, J. D. & Stickgold, R. The role of sleep in declarative memory consolidation: passive, permissive, active or none? Curr. Opin. Neurobiol. 16 , 716–722 (2006).
Payne, J. D. et al. Memory for semantically related and unrelated declarative information: the benefit of sleep, the cost of wake. PLoS One 7 , e33079 (2012).
Gaskell, M. G., Cairney, S. A. & Rodd, J. M. Contextual priming of word meanings is stabilized over sleep. Cognition 182 , 109–126 (2019).
Schabus, M. et al. Sleep spindles and their significance for declarative memory consolidation. Sleep 27 , 1479–1485 (2004).
Himmer, L., Müller, E., Gais, S. & Schönauer, M. Sleep-mediated memory consolidation depends on the level of integration at encoding. Neurobiol. Learn. Mem. 137 , 101–106 (2017).
Jacoby, L. L. A process dissociation framework: separating automatic from intentional uses of memory. J. Mem. Lang. 30 , 513–541 (1991).
Sigman, M. & Dehaene, S. Parsing a cognitive task: a characterization of the mind’s bottleneck. PLoS Biol. 3 , 0334–0349 (2005).
Cohen, D. A., Pascual-Leone, A., Press, D. Z. & Robertson, E. M. Off-line learning of motor skill memory: a double dissociation of goal and movement. Proc. Natl Acad. Sci. USA 102 , 18237–18241 (2005).
Conte, F. & Ficca, G. Caveats on psychological models of sleep and memory: a compass in an overgrown scenario. Sleep. Med. Rev. 17 , 105–121 (2013).
Stickgold, R. Parsing the role of sleep in memory processing. Curr. Opin. Neurobiol. 23 , 847–853 (2013).
Ackermann, S., Hartmann, F., Papassotiropoulos, A., De Quervain, D. J. F. & Rasch, B. No associations between interindividual differences in sleep parameters and episodic memory consolidation. Sleep 38 , 951–959 (2015).
Schendan, H. E., Searl, M. M., Melrose, R. J. & Stern, C. E. An fMRI study of the role of the medial temporal lobe in implicit and explicit sequence learning. Neuron 37 , 1013–1025 (2003).
Fischer, S., Drosopoulos, S., Tsen, J. & Born, J. Implicit learning–explicit knowing: a role for sleep in memory system interaction. J. Cogn. Neurosci. 18 , 311–319 (2006).
Simor, P. et al. Deconstructing procedural memory: different learning trajectories and consolidation of sequence and statistical learning. Front. Psychol. 9 , 2708 (2019).
Freedberg, M., Toader, A. C., Wassermann, E. M. & Voss, J. L. Competitive and cooperative interactions between medial temporal and striatal learning systems. Neuropsychologia 136 , 107257 (2020).
Poldrack, R. A., & Packard, M. G. Competition among multiple memory systems: converging evidence from animal and human brain studies. Neuropsychologia 41 , 245–251 (2003).
Brown, R. M. & Robertson, E. M. Off-line processing: reciprocal interactions between declarative and procedural memories. J. Neurosci. 27 , 10468–10475 (2007).
Viczko, J., Sergeeva, V., Ray, L. B., Owen, A. M. & Fogel, S. M. Does sleep facilitate the consolidation of allocentric or egocentric representations of implicitly learned visual-motor sequence learning? Learn. Mem. 25 , 67–77 (2018).
Albouy, G. et al. Daytime sleep enhances consolidation of the spatial but not motoric representation of motor sequence memory. PLoS One 8 , e52805 (2013).
Song, S. & Cohen, L. G. Practice and sleep form different aspects of skill. Nat. Commun. 5 , 3407 (2014).
Chan, J. C. K. & McDermott, K. B. The testing effect in recognition memory: a dual process account. J. Exp. Psychol. Learn. Mem. Cogn. 33 , 431–437 (2007).
Soderstrom, N. C., Kerr, T. K. & Bjork, R. A. The critical importance of retrieval — and spacing — for learning. Psychol. Sci. 27 , 223–230 (2016).
Karpicke, J. D. & Roediger, H. L. The critical importance of retrieval for learning. Science 319 , 966–968 (2008).
Wiklund-Hörnqvist, C., Jonsson, B. & Nyberg, L. Strengthening concept learning by repeated testing. Scand. J. Psychol. 55 , 10–16 (2014).
Buschke, H. Selective reminding for analysis of memory and learning. J. Verbal Learn. Verbal Behav. 12 , 543–550 (1973).
Quan, S. F., Budhiraja, R. & Kushida, C. A. Associations between sleep quality, sleep architecture and sleep disordered breathing and memory after continuous positive airway pressure in patients with obstructive sleep apnea in the Apnea Positive Pressure Long-term Efficacy Study (APPLES). Sleep. Sci. 11 , 231–238 (2018).
Uguccioni, G. et al. Sleep-related declarative memory consolidation and verbal replay during sleep talking in patients with REM sleep behavior disorder. PLoS ONE 8 , e83352 (2013).
Mazza, S. et al. Relearn faster and retain longer: along with practice, sleep makes perfect. Psychol. Sci. 27 , 1321–1330 (2016).
Ellenbogen, J. M., Hulbert, J. C., Stickgold, R., Dinges, D. F. & Thompson-Schill, S. L. Interfering with theories of sleep and memory: sleep, declarative memory, and associative interference. Curr. Biol. 16 , 1290–1294 (2006).
Pöhlchen, D., Pawlizki, A., Gais, S. & Schönauer, M. Evidence against a large effect of sleep in protecting verbal memories from interference. J. Sleep. Res. 30 , e13042 (2021).
Klinzing, J. G., Rasch, B., Born, J. & Diekelmann, S. Sleep’s role in the reconsolidation of declarative memories. Neurobiol. Learn. Mem. 136 , 166–173 (2016).
Fowler, M. J., Sullivan, M. J. & Ekstrand, B. R. Sleep and memory. Science 179 , 302–304 (1973).
Cordi, M. J., Diekelmann, S., Born, J. & Rasch, B. No effect of odor-induced memory reactivation during REM sleep on declarative memory stability. Front. Syst. Neurosci. 8 , 100576 (2014).
Prehn-Kristensen, A. et al. Transcranial oscillatory direct current stimulation during sleep improves declarative memory consolidation in children with attention-deficit/hyperactivity disorder to a level comparable to healthy controls. Brain Stimul. 7 , 793–799 (2014).
Fenn, K. M. & Hambrick, D. Z. Individual differences in working memory capacity predict sleep-dependent memory consolidation. J. Exp. Psychol. Gen. 141 , 404–410 (2012).
Wilhelm, I., Diekelmann, S. & Born, J. Sleep in children improves memory performance on declarative but not procedural tasks. Learn. Mem. 15 , 373–377 (2008).
Marshall, L., Mölle, M., Hallschmid, M. & Born, J. Transcranial direct current stimulation during sleep improves declarative memory. J. Neurosci. 24 , 9985–9992 (2004).
Xue, G. et al. Greater neural pattern similarity across repetitions is associated with better memory. Science 330 , 97–101 (2010).
Young, D. R. & Bellezza, F. S. Encoding variability, memory organization, and the repetition effect. J. Exp. Psychol. Learn. Mem. Cogn. 8 , 545–559 (1982).
Ebbinghaus, H. Memory: a contribution to experimental psychology. Ann. Neurosci. 20 , 155 (2013).
Maski, K. et al. Sleep dependent memory consolidation in children with autism spectrum disorder. Sleep 38 , 1955–1963 (2015).
Baran, B. et al. Spared and impaired sleep-dependent memory consolidation in schizophrenia. Schizophr. Res. 199 , 83–89 (2018).
Nissen, M. J. & Bullemer, P. Attentional requirements of learning: evidence from performance measures. Cogn. Psychol. 19 , 1–32 (1987).
Brawn, T. P., Fenn, K. M., Nusbaum, H. C. & Margoliash, D. Consolidating the effects of waking and sleep on motor-sequence learning. J. Neurosci. 30 , 13977–13982 (2010).
Török, B., Janacsek, K., Nagy, D. G., Orbán, G. & Nemeth, D. Measuring and filtering reactive inhibition is essential for assessing serial decision making and learning. J. Exp. Psychol. Gen. 146 , 529–542 (2017).
Rickard, T. C., Cai, D. J., Rieth, C. A., Jones, J. & Ard, M. C. Sleep does not enhance motor sequence learning. J. Exp. Psychol. Learn. Mem. Cogn. 34 , 834–842 (2008).
Cai, D. J. & Rickard, T. C. Reconsidering the role of sleep for motor memory. Behav. Neurosci. 123 , 1153–1157 (2009).
Nettersheim, A., Hallschmid, M., Born, J. & Diekelmann, S. The role of sleep in motor sequence consolidation: stabilization rather than enhancement. J. Neurosci. 35 , 6696–6702 (2015).
Rieth, C. A., Cai, D. J., McDevitt, E. A. & Mednick, S. C. The role of sleep and practice in implicit and explicit motor learning. Behav. Brain Res. 214 , 470–474 (2010).
Rickard, T. C., Pan, S. C. & Gupta, M. W. Severe publication bias contributes to illusory sleep consolidation in the motor sequence learning literature. J. Exp. Psychol. Learn. Mem. Cogn. 48 , 1787–1796 (2022).
Ioannidis, J. P. A. Why most published research findings are false. PLoS Med. 2 , e124 (2005).
Maxwell, S. E., Lau, M. Y. & Howard, G. S. Is psychology suffering from a replication crisis?: What does ‘failure to replicate’ really mean? Am. Psychol. 70 , 487–498 (2015).
Aarts, A. A. et al. Estimating the reproducibility of psychological science. Science 349 , aac4716 (2015).
Wagner, U., Hallschmid, M., Rasch, B. & Born, J. Brief sleep after learning keeps emotional memories alive for years. Biol. Psychiat. 60 , 788–790 (2006).
Csábi, E. et al. Declarative and non-declarative memory consolidation in children with sleep disorder. Front. Hum. Neurosci. 9 , 709 (2016).
Newbury, C. R., Crowley, R., Rastle, K. & Tamminen, J. Sleep deprivation and memory: meta-analytic reviews of studies on sleep deprivation before and after learning. Psychol. Bull. 147 , 1215–1240 (2021).
Serdar, C. C., Cihan, M., Yücel, D. & Serdar, M. A. Sample size, power and effect size revisited: simplified and practical approachin pre-clinical, clinical and laboratory studies. Biochem. Med. 31 , 010502 (2021).
Berres, S. & Erdfelder, E. The sleep benefit in episodic memory: an integrative review and a meta-analysis. Psychol. Bull. 147 , 1309–1353 (2021).
Wagenmakers, E.-J., Lee, M., Lodewyckx, T. & Iverson, G. J. in Bayesian Evaluation of Informative Hypotheses 181–207 (Springer, 2008).
American Psychological Association. (2010). Publication manual of the American psychological association. American Psychological Association.
Dienes, Z. How Bayes factors change scientific practice. J. Math. Psychol. 72 , 78–89 (2016).
Article MathSciNet Google Scholar
Dienes, Z., Coulton, S. & Heather, N. Using Bayes factors to evaluate evidence for no effect: examples from the SIPS project. Addiction 113 , 240–246 (2018).
Dienes, Z. & Mclatchie, N. Four reasons to prefer Bayesian analyses over significance testing. Psychon. Bull. Rev. 25 , 207–218 (2018).
Wagenmakers, E. J. et al. Bayesian inference for psychology. Part I: Theoretical advantages and practical ramifications. Psychon. Bull. Rev. 25 , 35–57 (2018).
Rouder, J. N. Optional stopping: no problem for Bayesians. Psychon. Bull. Rev. 21 , 301–308 (2014).
Schönbrodt, F. D., Wagenmakers, E. J., Zehetleitner, M. & Perugini, M. Sequential hypothesis testing with Bayes factors: efficiently testing mean differences. Psychol. Meth. 22 , 322–339 (2017).
Brown, H. & Maylor, E. A. Memory consolidation effects on memory stabilization and item integration in older adults. Psychon. Bull. Rev. 24 , 1032–1039 (2017).
Van De Schoot, R., Winter, S. D., Ryan, O., Zondervan-Zwijnenburg, M. & Depaoli, S. Supplemental material for a systematic review of Bayesian articles in psychology: the last 25 years. Psychol. Meth. 22 , 217–239 (2017).
Ellis, P. D. The Essential Guide to Effect Sizes: Statistical Power, Meta-Analysis, and the Interpretation of Research Results (Cambridge Univ. Press, 2010).
Scullin, M. K. Sleep, memory, and aging: the link between slow-wave sleep and episodic memory changes from younger to older adults. Psychol. Aging 28 , 105–114 (2013).
Ujma, P. P. Sleep spindles and general cognitive ability — a meta-analysis. Sleep. Spindl. Cort. Up States 2 , 1–17 (2018).
Google Scholar
Sturman, M. C., Sturman, A. J. & Sturman, C. J. Uncontrolled control variables: the extent that a researcher’s degrees of freedom with control variables increases various types of statistical errors. J. Appl. Psychol. 107 , 9–22 (2022).
Lerner, I., Lupkin, S. M., Tsai, A., Khawaja, A. & Gluck, M. A. Sleep to remember, sleep to forget: rapid eye movement sleep can have inverse effects on recall and generalization of fear memories. Neurobiol. Learn. Mem. 180 , 107413 (2021).
Abdi, H. in Encyclopedia of Measurement and Statistics (ed. Salkind, N. J.) 103–107 https://doi.org/10.4135/9781412952644 (Sage, 2007).
Forstmeier, W., Wagenmakers, E. J. & Parker, T. H. Detecting and avoiding likely false-positive findings — a practical guide. Biol. Rev. 92 , 1941–1968 (2017).
Kruschke, J. K. Bayesian analysis reporting guidelines. Nat. Hum. Behav. 5 , 1282–1291 (2021).
Bódizs, R. et al. Prediction of general mental ability based on neural oscillation measures of sleep. J. Sleep. Res. 14 , 285–292 (2005).
Fang, Z. et al. Sleep spindles and intellectual ability: epiphenomenon or directly related? J. Cogn. Neurosci. 29 , 167–182 (2017).
Fang, Z., Ray, L. B., Owen, A. M. & Fogel, S. M. Brain activation time-locked to sleep spindles associated with human cognitive abilities. Front. Neurosci. 13 , 432131 (2019).
Fogel, S. M. & Smith, C. T. Learning-dependent changes in sleep spindles and stage 2 sleep. J. Sleep. Res. 15 , 250–255 (2006).
Daw, N. D., Niv, Y. & Dayan, P. Uncertainty-based competition between prefrontal and dorsolateral striatal systems for behavioral control. Nat. Neurosci. 8 , 1704–1711 (2005).
Fogel, S. M. & Smith, C. T. The function of the sleep spindle: a physiological index of intelligence and a mechanism for sleep-dependent memory consolidation. Neurosci. Biobehav. Rev. 35 , 1154–1165 (2011).
Lahl, O., Wispel, C., Willigens, B. & Pietrowsky, R. An ultra short episode of sleep is sufficient to promote declarative memory performance. J. Sleep. Res. 17 , 3–10 (2008).
Diekelmann, S., Biggel, S., Rasch, B. & Born, J. Offline consolidation of memory varies with time in slow wave sleep and can be accelerated by cuing memory reactivations. Neurobiol. Learn. Mem. 98 , 103–111 (2012).
Nishida, M. & Walker, M. P. Daytime naps, motor memory consolidation and regionally specific sleep spindles. PLoS One 2 , e341 (2007).
Schönauer, M., Geisler, T. & Gais, S. Strengthening procedural memories by reactivation in sleep. J. Cogn. Neurosci. 26 , 143–153 (2014).
Gais, S., Plihal, W., Wagner, U. & Born, J. Early sleep triggers memory for early visual discrimination skills. Nat. Neurosci. 3 , 1335–1339 (2000).
Mednick, S. C., Cai, D. J., Kanady, J. & Drummond, S. P. A. Comparing the benefits of caffeine, naps and placebo on verbal, motor and perceptual memory. Behav. Brain Res. 193 , 79–86 (2008).
Sopp, M. R., Michael, T. & Mecklinger, A. Effects of early morning nap sleep on associative memory for neutral and emotional stimuli. Brain Res. 1698 , 29–42 (2018).
Plihal, W., Weaver, S., Mölle, M., Fehm, H. L. & Born, J. Sensory processing during early and late nocturnal sleep. Electroencephalogr. Clin. Neurophysiol. 99 , 247–256 (1996).
Yaroush, R., Sullivan, M. J. & Ekstrand, B. R. Effect of sleep on memory: II. Differential effect of the first and second half of the night. J. Exp. Psychol. 88 , 361–366 (1971).
Mednick, S., Nakayama, K. & Stickgold, R. Sleep-dependent learning: a nap is as good as a night. Nat. Neurosci. 6 , 697–698 (2003).
Wamsley, E. J. Memory consolidation during waking rest. Trends Cogn. Sci. 23 , 171–173 (2019).
McDevitt, E. A., Duggan, K. A. & Mednick, S. C. REM sleep rescues learning from interference. Neurobiol. Learn. Mem. 122 , 51–62 (2015).
Cousins, J. N., Wong, K. F., Raghunath, B. L., Look, C. & Chee, M. W. L. The long-term memory benefits of a daytime nap compared with cramming. Sleep 42 , zsy207 (2019).
Piosczyk, H. et al. The effect of sleep-specific brain activity versus reduced stimulus interference on declarative memory consolidation. J. Sleep. Res. 22 , 406–413 (2013).
Schichl, M., Ziberi, M., Lahl, O. & Pietrowsky, R. The influence of midday naps and relaxation-hypnosis on declarative and procedural memory performance. Sleep. Hypn. 13 , 7–14 (2011).
Mednick, S. C., Makovski, T., Cai, D. J. & Jiang, Y. V. Sleep and rest facilitate implicit memory in a visual search task. Vis. Res. 49 , 2557–2565 (2009).
Schapiro, A. C., McDevitt, E. A., Rogers, T. T., Mednick, S. C. & Norman, K. A. Human hippocampal replay during rest prioritizes weakly learned information and predicts memory performance. Nat. Commun. 9 , 3920 (2018).
van Schalkwijk, F. J. et al. The effect of daytime napping and full‐night sleep on the consolidation of declarative and procedural information. J. Sleep Res. 28 , e12649 (2019).
Poldrack, R. A. & Gorgolewski, K. J. OpenfMRI: open sharing of task fMRI data. Neuroimage 144 , 259–261 (2017).
Gorgolewski, K., Esteban, O., Schaefer, G., Wandell, B. & Poldrack, R. OpenNeuro — a free online platform for sharing and analysis of neuroimaging data. F1000Res. 6 , 80190187 (2017).
Spitschan, M., Schmidt, M. H. & Blume, C. Principles of open, transparent and reproducible science in author guidelines of sleep research and chronobiology journals. Wellcome Open Res . https://wellcomeopenresearch.org/articles/5-172 (2021).
Prabhakar, J., Johnson, E. G., Nordahl, C. W. & Ghetti, S. Memory-related hippocampal activation in the sleeping toddler. Proc. Natl Acad. Sci. USA 115 , 6500–6505 (2018).
Schreiner, T., Petzka, M., Staudigl, T. & Staresina, B. P. Endogenous memory reactivation during sleep in humans is clocked by slow oscillation-spindle complexes. Nat. Commun. 12 , 3112 (2021).
Rakowska, M., Abdellahi, M. E. A., Bagrowska, P., Navarrete, M. & Lewis, P. A. Long term effects of cueing procedural memory reactivation during NREM sleep. Neuroimage 244 , 118573 (2021).
Nosek, B. A. et al. Promoting an open research culture. Science 348 , 1422–1425 (2015).
Simor, P., Gombos, F., Blaskovich, B. & Bódizs, R. Long-range alpha and beta and short-range gamma EEG synchronization distinguishes phasic and tonic REM periods. Sleep https://doi.org/10.1093/sleep/zsx210 (2018).
Simor, P., van Der Wijk, G., Gombos, F. & Kovács, I. The paradox of rapid eye movement sleep in the light of oscillatory activity and cortical synchronization during phasic and tonic microstates. Neuroimage 202 , 116066 (2019).
Foster, E. D. & Deardorff, A. Open science framework (OSF). J. Med. Libr. Assoc. 105 , 203–206 (2017).
Article PubMed Central Google Scholar
Nosek, B. A., Ebersole, C. R., DeHaven, A. C. & Mellor, D. T. The preregistration revolution. Proc. Natl Acad. Sci. USA 115 , 2600–2606 (2018).
Lindsay, D. S., Simons, D. J. & Lilienfeld, S. O. Research preregistration 101. APS https://www.psychologicalscience.org/observer/research-preregistration-101 (2016).
Hardwicke, T. E. & Wagenmakers, E. J. Reducing bias, increasing transparency and calibrating confidence with preregistration. Nat. Hum. Behav. 7 , 15–26 (2023).
Angelis, C. D. et al. Clinical trial registration: a statement from the International Committee of Medical Journal Editors. Lancet 364 , 911–912 (2004).
James, E., Gaskell, M. G. & Henderson, L. M. Offline consolidation supersedes prior knowledge benefits in children’s (but not adults’) word learning. Dev. Sci. 22 , e12776 (2019).
Soderberg, C. K. et al. Initial evidence of research quality of registered reports compared with the standard publishing model. Nat. Hum. Behav. 5 , 990–997 (2021).
Download references
Acknowledgements
The authors thank K. Schipper for her help and comments on the manuscript. This research was supported by the ANR grant awarded within the framework of the Inserm CPJ ANR-22-CPJ1-0042-01 (to D.N.); Hungary’s National Brain Research Program (project NAP2022-I-2/2022); NKFIH-OTKA PD 124148 (Principal Investigator K.J.); NKFI FK 142945 (Principal Investigator P.S.); Janos Bolyai Research Fellowship of the Hungarian Academy of Sciences (to K.J. and P.S.); and the French National Agency for Research (ANR, grant number ANR-15-CE33-0003, Principal Investigator S.M.).
Author information
These authors contributed equally: Dezső Németh, Emilie Gerbier, Karolina Janacsek.
Authors and Affiliations
Centre de Recherche en Neurosciences de Lyon CRNL U1028 UMR5292, INSERM, CNRS, Université Claude Bernard Lyon, Bron, France
Dezső Németh, Perrine Ruby & Teodóra Vékony
Institute of Psychology, ELTE Eötvös Lorand University, Budapest, Hungary
Dezső Németh, Peter Simor & Karolina Janacsek
Institute of Cognitive Neuroscience and Psychology, Hun-Ren Research Centre for Natural Sciences, Budapest, Hungary
Dezső Németh
Laboratoire Bases, Corpus, Langage (UMR 7320), CNRS, Université Côte d’Azur, Nice, France
Emilie Gerbier
Institute of Medical Psychology and Behavioral Neurobiology, University of Tübingen, Tübingen, Germany
Jan Born & Susanne Diekelmann
Department of Psychology, University of California San Diego, La Jolla, CA, USA
Timothy Rickard
Department of Psychiatry and Psychotherapy, University Hospital Tübingen, Tübingen, Germany
Susanne Diekelmann
School of Psychology, University of Ottawa, Ottawa, Canada
Stuart Fogel
Donders Institute for Brain Cognition and Behaviour, Radboud University, Nijmegen, Netherlands
Lisa Genzel
Institute for Child and Adolescent Psychiatry, Center for Integrative Psychiatry, University Hospital Schleswig-Holstein, Kiel, Germany
Alexander Prehn-Kristensen
Department of Psychology, Faculty of Human Sciences, MSH Medical School Hamburg, University of Applied Sciences and Medical University, Hamburg, Germany
Department of Psychology, University of Notre Dame, Notre Dame, IN, USA
Jessica Payne
Donders Institute for Brain, Cognition and Behavior, Radboud University Medical Center, Nijmegen, The Netherlands
Martin Dresler
Institute of Behavioural Sciences, Semmelweis University, Budapest, Hungary
Peter Simor
Forgetting Team, Centre de Recherche en Neurosciences de Lyon CRNL U1028 UMR5292, INSERM, CNRS, Université Claude Bernard Lyon, Bron, France
Stephanie Mazza
Department of Psychology, Centre for Cognitive Neuroscience, University of Salzburg, Salzburg, Austria
Kerstin Hoedlmoser & Manuel Schabus
Department of Psychological and Brain Sciences, University of Massachusetts, Amherst, MA, USA
Rebecca M. C. Spencer
Department of Movement Sciences, Katholieke Universiteit Leuven, Leuven, Belgium
Genevieve Albouy
Centre for Thinking and Learning, Institute for Lifecourse Development, School of Human Sciences, University of Greenwich, London, UK
Karolina Janacsek
You can also search for this author in PubMed Google Scholar
Contributions
D.N., E.G., J.B., T.R., S.D., S.F., L.G., A.P.-K., J.P., M.D., P.S., S.M., K.H., P.R., R.M.C.S., G.A., T.V., M.S. and K.J. contributed substantially to discussion of the content. All authors wrote the article. All authors reviewed and/or edited the manuscript before submission.
Corresponding author
Correspondence to Dezső Németh .
Ethics declarations
Competing interests.
The authors declare no competing interests.
Peer review
Peer review information.
Nature Reviews Psychology thanks M. Gareth Gaskell and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
Reprints and permissions
About this article
Cite this article.
Németh, D., Gerbier, E., Born, J. et al. Optimizing the methodology of human sleep and memory research. Nat Rev Psychol 3 , 123–137 (2024). https://doi.org/10.1038/s44159-023-00262-0
Download citation
Published : 22 December 2023
Issue Date : February 2024
DOI : https://doi.org/10.1038/s44159-023-00262-0
Share this article
Anyone you share the following link with will be able to read this content:
Sorry, a shareable link is not currently available for this article.
Provided by the Springer Nature SharedIt content-sharing initiative
Quick links
- Explore articles by subject
- Guide to authors
- Editorial policies
Sign up for the Nature Briefing newsletter — what matters in science, free to your inbox daily.

Memory and cognition: shifting approaches to how we do science
- Published: 16 February 2023
- Volume 51 , pages 1–3, ( 2023 )
Cite this article
- Ayanna K. Thomas 1
2235 Accesses
2 Altmetric
Explore all metrics
Avoid common mistakes on your manuscript.
On 1 January 2020, I took on the role as Editor-in-Chief of Memory and Cognition . My vision has been and continues to be to create a venue for research that broadens our understanding of human cognition. I seek to do this by encouraging more research that examines cognition in context and expands the diversity of the included content area. I continue to support the publication of innovative methodological and analytical perspectives. I strive to develop and offer tools to foster visibility of new ideas. And I aim to improve the representation of culture, demography, and geography of readers, authors, reviewers, and the editorial staff. With the start of 2023, I not only remind our readership of what we strive for at Memory and Cognition , but also offer some reflection on what I have learned in these 3 years as Editor. I offer this editorial as a glimpse into how the commitment to support theory-driven research, embrace fresh perspectives, encourage diversity of thought, and enable the team of scientists who serve as associate editors for this journal have evolved.
The year 2020 brought a unique set of challenges for our world. With a global pandemic that forced us all into isolation, the team at Memory and Cognition had to quickly pivot in how we approached the process of reviewing manuscripts. We carefully considered procedures to reduce unnecessary burdens on reviewers and developed strategies to make timely, effective, and efficient decisions. We found that, even under the pressures of isolation and illness, many in the scientific community were willing to dedicate time and energy to the peer-review process. 2020 emerged as an exceptionally impactful year for Memory and Cognition . We accepted approximately 150 empirical articles presenting innovative research spanning visual statistical learning to analogical reasoning. In addition, we developed a special issue focused on reconceptualizing the distinction between episodic and semantic memory edited by Felipe De Brigard, Muireann Irish, and Sharda Umanath. I am proud of the work that our community produced during that year, and I am especially impressed by the Associate Editors (Sarah Barber, Monica Bucciarelli, Jennifer Coane, Felipe De Brigard, Bridgid Finn, Wendy Francis, Joe Magliano, Steve Majerus, Henry Otgaar, Priti Shah, Laura Thomas) and every reviewer who devoted time that year.
However, 2020 brought more than the disruption and tragedy of a pandemic that still costs lives daily. On 25 May 2020, George Perry Floyd Jr. was murdered by police officers in Minneapolis, Minnesota. Although the tragedy of George Floyd’s murder is one that is all too familiar in the US landscape, this event sparked a movement across the USA that centered critical discussions of race, racism, and anti-racism. This movement directly impacted our field and shaped my thinking on not only whose voices are included in scientific discussions, but also what questions are valued for research.
The way we are trained to think about human cognition and the way in which we evaluate others’ work has subtly limited advancing our understanding of the variables that affect our thinking and behavior. I and others have argued that the current predominant approaches to study have limited reliable theory development, methodological advances, and understanding of the function and processes that underlie human cognition (e.g., Prather, 2021 ; Thomas et al., in press ).
The research presented in this journal since its inception has focused squarely on the understanding of human cognition as a set of internal processes that allow for the transformation of sensory input into abstract mental representations that can be stored, recovered, and used. The predominant approach to the study of human cognition is on understanding internal cognitive processes, informed by the assumption that cognitive processes are generally universal across the human species (Thomas et al., in press ). The pursuit of cognitive universals has led to explicitly associating the brain and biology to cognition, and presupposes that beneath superficial differences that may emerge across cultures, or deficits in performance that may emerge across racial groups, lies a shared cognitive architecture (Block, 1995 ). It is from this perspective that the pursuit of universals was an important counterpoint to the scientific racism that preceded and continues to co-exist with the field of Cognitive Psychology. However, the era of objective, context-free, empirical investigation into human cognition did not ameliorate the persistence of scientific racism. Rather, the context-free approach taken by many cognitive researchers in the pursuit of universal cognitive processes may have limited our ability as scientists to effectively characterize human cognition.
The discussion of race in the history of Cognitive Psychology is not new. However, we as a field are becoming increasingly aware that structural racism has impacted our characterization of “normal” or “control” participant groups. We are beginning to recognize that principles of human cognition may not be effectively derived from unrepresentative samples (e.g., WEIRD populations), that questions regarding human cognition may not be effectively generated from a homogenous set of scientists, that human cognition may be better understood in context, and that individual and cultural variation should be investigated and leveraged to support robust theories of human cognition (Prather, 2021 ).
Recognizing the need for diverse samples, ecologically relevant stimuli, and considering context in understanding cognition has led to important findings in categorization (e.g., Medin & Atran, 2004 ), perception (e.g., Haddon et al., 2011 ; Segall et al., 1963 ), language development (Figueroa, in press ), and memory (Wang, 2021 ), to name a few. As one example, research has shown that Westerners are more likely to attend to and encode object information independent of context as compared to East Asian individuals. This difference in encoding suggests perceptual and cognitive processes that align with cultural orientations, practices, and ecologies. Ignoring these influences or establishing norms of cognitive processes based on restricted cultural perspectives limits the utility of our findings. Considering new or understudied approaches to cognition may lead us to embrace cognition more fully as a complex system composed of many internal individual elements embedded within, and open to an external complex environment. We may begin to consider human cognition as enduring patterns of reactions, attitudes, or overt behavioral manifestations that are a function of demographic, cultural, social, and other environmental factors (Boykin, 1977 ).
In the 2 years I have remaining as Editor-in-Chief of this journal, I would like to create a space for investigations and discussions that consider cognition in context. We have a forthcoming special issue focusing on exploring the assumptions of cognitive universality and human cognitive diversity to be published in 2023. In addition, Memory and Cognition will offer a venue for broadening the field’s perspective on the study of human cognition. To achieve this goal, Memory and Cognition will now accept new formats for publications.
Types of New Articles Published
Commentary Papers and Review Articles that
Introduce new theoretical ideas, methods, and/or approaches to studying cognitive psychology.
Map basic cognitive psychology research onto applied and translational science.
Discuss methods and practices for engaging citizens in our scientific pursuits.
Discussion papers that debate or discuss a specific topic in Cognitive Psychology from multiple perspectives.
Special Issues and Manuscripts that focus on
Examining cognition in context.
Combining complementary theories and/or findings from distinct research traditions.
Memory and Cognition will continue to accept empirical quantitative research that examines human memory and learning, conceptual processes, psycholinguistics, problem solving, thinking, decision making, and skilled performance. We will continue to review relevant work in the areas of computer simulation, information processing, mathematical psychology, developmental psychology, and experimental social psychology. The core principles of Memory and Cognition remain central to this vision. The goal of these new article formats is to encourage novel approaches to theory and methods that embrace examining problems from multiple angles and conflicting perspectives.
As Editor, I continue to champion the diversity of research questions (e.g., interdisciplinary and inter-sub-disciplinary questions) and approaches (e.g., computational, individual differences) published in Memory and Cognition . I encourage the reader to take this opportunity to consider this journal as a venue that upholds the traditions of theoretically robust empirical research while embracing new methods and perspectives that allow for a broader consideration of complexity in cognition.
Although I end my tenure as Editor-in-Chief at the end of 2024, I hope to encourage a new era at Memory and Cognition that broadens the way we think about human cognition.
Block, N. (1995). The mind as the software of the brain. In Thinking: An invitation to cognitive science (Vol. 3, 2nd ed., pp. 377–425). The MIT Press.
Google Scholar
Boykin, A. W. (1977). Experimental psychology from a black perspective: Issues and examples. Journal of Black Psychology, 3 (2), 29–49.
Article Google Scholar
Figueroa, M. (in press). Language development, linguistic input, and linguistic racism . Wires Cognitive Science.
Haddon, A. C., Rivers, W. H. R, & Wilkins, A. (2011). Reports of the Cambridge Anthropological Expedition to Torres Straits . Cambridge University Press.
Medin, D. L., & Atran, S. (2004). The native mind: Biological categorization and reasoning in development and across cultures. Psychological Review, 111 (4), 960–983. https://doi.org/10.1037/0033-295X.111.4.960
Article PubMed Google Scholar
Prather, R. (2021). Reconstructing the Study of Human Cognitio . https://doi.org/10.31234/osf.io/45a2q
Segall, M. H., Campbell, D. T., & Herskovits, M. J. (1963). Cultural differences in the perception of geometric illusions. Science . https://doi.org/10.1126/science.139.3556.769
Thomas, A. K., McKinney de Royston, M., & Powell, S. (in press). Color evasive cognition: The unavoidable impact of scientific racism in the founding of a field. Current Directions in Psychological Science .
Wang, Q. (2021). The Cultural Foundation of human memory. Annual Review of Psychology, 72 , 151–179. https://doi.org/10.1146/annurev-psych-070920-023638
Download references
Author information
Authors and affiliations.
Tufts University, Psychology, Medford, MA, USA
Ayanna K. Thomas
You can also search for this author in PubMed Google Scholar
Corresponding author
Correspondence to Ayanna K. Thomas .
Additional information
Publisher’s note.
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Reprints and permissions
About this article
Thomas, A.K. Memory and cognition: shifting approaches to how we do science. Mem Cogn 51 , 1–3 (2023). https://doi.org/10.3758/s13421-023-01395-6
Download citation
Accepted : 10 January 2023
Published : 16 February 2023
Issue Date : January 2023
DOI : https://doi.org/10.3758/s13421-023-01395-6
Share this article
Anyone you share the following link with will be able to read this content:
Sorry, a shareable link is not currently available for this article.
Provided by the Springer Nature SharedIt content-sharing initiative
- Find a journal
- Publish with us
- Track your research
Colorado State University
Human Memory Lab

Research Interests
In this lab, we study human memory. Though we study a broad range of issues related to human memory processes and performance, the primary research interest in this lab is in people’s ability to show memory when they experience retrieval failure. For example, people sometimes fail to recall the information that they are searching for in memory. When experiencing such memory failure, people often still have other information available to them about the memory—information that can at times be used to make decisions with some degree of accuracy. An example is the phenomenon recognition without identification and a seemingly similar phenomenon, recognition without recall . We investigate the various ways that people can show memory when recall fails, and also the neural mechanisms that underlie memory that occurs when retrieval fails. In this lab, we use many methodological techniques for addressing these questions, including behavioral paradigms, virtual reality technology, brain electrophysiological techniques, mathematical model simulations, and, in collaboration with other laboratories and facilities, functional magnetic resonance imaging (or fMRI) and research with clinical populations.
Basic Memory & Metacognition Research
Familiarity-based Recognition. One example of a form of memory that can occur in the face of retrieval failure is familiarity, which is the feeling of having experienced something before, without being able to pinpoint exactly why the feeling is occurring. One branch of research in this lab aims to study familiarity as a basis for recognition and how it operates mechanistically and how it might differ from other processes that enable recognition.
Features that Produce Recognition when Recall Fails. One branch of research in this lab attempts to identify what features of an item or situation can produce recognition when recall fails. For example, when retrieval fails, do geometric shapes contribute to a sense of recognition with pictures and objects? Do phonemes contribute to a sense of recognition with spoken words? Can rhythm contribute to a sense of recognition with songs? Can more abstract features, such as semantic features, produce a sense of recognition when retrieval fails?
Subjective Memory Experiences that Occur During Retrieval Failure. The ability to recognize having had a prior experience with a situation when recall of the exact prior experience itself fails is related to the subjective experience of sensing that something is in memory. Besides the feeling of familiarity, an example of one such type of subjective experience is the déjà vu experience, which occurs when one has a feeling of having experienced something before, despite evidence to the contrary. Most commonly, déjà vu occurs with places—people experience a feeling of having been somewhere before, despite knowing that they have never been there. Déjà vu may result from a memory that fails to be retrieved, as is humorously illustrated in this commercial . One branch of research in this lab aims to better understand déjà vu as a memory phenomenon, and virtual reality is one of the tools that we use in this lab to investigate this ( click here for a video description ). Another example of a type of subjective memory experience that occurs during retrieval failure is the tip-of-the-tongue (TOT) phenomenon, whereby one feels that a word is in memory, but cannot currently access it. One branch of research in this lab investigates the TOT phenomenon and how it differs from other seemingly similar subjective memory experiences that occur during retrieval failure. Another branch is aimed at investigating how TOT states affect other cognitive judgments during the uncertainty of retrieval failure.
Unconscious Recognition. Other ways that people can show memory when retrieval fails may be unconscious. At least some evidence from this lab and others suggests that people may sometimes be influenced by prior memories to choose certain items over others or to react in certain ways to situations, yet be completely unaware that situations in memory are driving those decisions. Another branch of research in our lab aims to study this ability and how it differs from conscious forms of memory that occur when retrieval fails.
Applied Memory & Metacognition Research
Applied Science of Learning Strategies for Reinforcing Learning Outside of the Classroom. One applied line of research in our lab focuses on investigating strategies for helping students to reinforce their learning outside of the classroom. For example: Wearable Technology for Automatizing Science-based Study Strategies: Reinforcing Learning Through Intermittent Smartwatch Prompting – ScienceDirect
Investigating how Subjective Metacognitive Sensations of Memory Might be Applicable to Learning & Training Situations. One applied line of research in our lab focuses on how subjective metacognitive sensations of memory, like tip-of-the-tongue states, might sometimes be useful in educational and training settings. For example: The tip-of-the-tongue state as a form of access to information: Use of tip-of-the-tongue states for strategic adaptive test-taking – PubMed (nih.gov)

Representative Publications
- Cleary, A. M., Irving, Z. C., & Mills, C. (2023). What Flips Attention? Cognitive Science , 47 (4), e13274. https://doi.org/10.1111/cogs.13274
- Huebert, A. M., McNeely-White, K. L., & Cleary, A. M. (2023). On the relationship between tip-of-the-tongue states and partial recollective experience: Illusory partial recollective access during tip-of-the-tongue states. Journal of Experimental Psychology: General. Advance online publication. https://doi.org/10.1037/xge0001292
- Neisser, J., Abreu, G., Drane, D.L., Pedersen, N.P., Parsons, T.D., & Cleary, A.M. (2023). Opening a conceptual space for metamemory experience. New Ideas in Psychology , 69 , 100995. https://doi.org/10.1016/j.newideapsych.2022.100995
- Carlaw, B.N., Huebert, A.M., McNeely-White, K.L. et al. (2022). Detecting a familiar person behind the surgical mask: recognition without identification among masked versus sunglasses-covered faces. Cogn. Research 7 , 90. https://doi.org/10.1186/s41235-022-00440-3
- Huebert, A. M., & Cleary, A. M. (2022). Do first and last letters carry more weight in the mechanism behind word familiarity? Psychonomic Bulletin & Review , 1–8. Advance online publication. https://doi.org/10.3758/s13423-022-02093-1
- Huebert, A. M., McNeely-White, K. L., & Cleary, A. M. (2022). Can cue familiarity during recall failure prompt illusory recollective experience? Memory & Cognition , 50 (4), 681–695. https://doi.org/10.3758/s13421-021-01248-0
- McNeely-White, K.L., McNeely-White, D.G., Huebert, A.M., Carlaw, B.N., & Cleary, A.M. (2022). Specifying a relationship between semantic and episodic memory in the computation of a feature-based familiarity signal using MINERVA 2. Memory & Cognition. https://doi.org/10.3758/s13421-021-01234-6
- Cleary, A.M., Neisser, J., McMahan, T., Parsons, T.D., Alwaki, A., Okada, N., Vosoughi, A., Kheder, A., Drane, D.L., & Pedersen, N.P. (2021). Subjective distinguishability of seizure and non-seizure deja vu: A case report, brief literature review, and research prospects. Epilepsy & Behavior. 10.1016/j.yebeh.2021.108373
- Cleary, A.M., McNeely-White, K.L., Hausman, H., Dawson, J., Kuhn, S., Osborn, R.M., Huebert, A.M., & Rhodes, M.G. (2021). Wearable technology for automatizing science-based study strategies: Reinforcing learning through intermittent smartwatch prompting. Journal of Applied Research in Memory and Cognition. https://doi.org/10.1016/j.jarmac.2021.01.001
- Ryals, A.J., Kelly, M.E., & Cleary, A.M. (2021). Increased pupil dilation during tip-of-the-tongue states. Consciousness and Cognition, 92, 103152. https://doi.org/10.1016/j.concog.2021.103152
- McNeely-White, K.L., McNeely-White, D.G., & Cleary, A.M. (2021). Global matching in music familiarity: How musical features combine across memory traces to increase familiarity with the whole in which they are embedded. Journal of Memory and Language, 118, 104217. https://doi.org/10.1016/j.jml.2020.104217
- Cleary, A.M., McNeely-White, K.L., Russell, S.A., Huebert, A.M., & Hausman, H. (2021). The tip-of-the-tongue state as a form of access to information: Use of tip-of-the-tongue states for strategic adaptive test-taking. Journal of Applied Research in Memory and Cognition, 10, 131-142. https://doi.org/10.1016/j.jarmac.2020.08.013
- Cleary, A.M., Huebert, A.M., & McNeely-White, K.L. (2020). The deja vu phenomenon’s entry into the realm of science. In Cleary, A.M. & Schwartz, B.L. (Eds.). Memory Quirks: The Study of Odd Phenomena in Memory. Routledge. (pp. 271-287).
- Cleary, A.M., Huebert, A.M., McNeely-White, K.L., Spahr, K.S. (2019). A postdictive bias associated with deja vu. Psychonomic Bulletin & Review, 26, 1433-1439.
- McNeely-White, K.L., & Cleary, A.M. (2019). Music recognition without identification and its relation to deja entendu: A study using “Piano Puzzlers.” New Ideas in Psychology, 55, 50-57.
- Cleary, A.M., & Claxton, A.B. (2018). Deja vu: An illusion of prediction. Psychological Science, 29, 635-644.
- Cleary, A.M., Ryals, A.J., & Wagner, S.M (2016). Recognition during recall failure: Semantic feature matching as a mechanism for recognition of semantic cues when recall fails. Memory & Cognition, 44, 50-62.
- Cleary, A.M. & Claxton, A.B. (2015). The tip-of-the-tongue heuristic: How tip-of-the-tongue states confer perceptibility on inaccessible words. Journal of Experimental Psychology: Learning, Memory, and Cognition, 41, 1533-1539.
- Cleary, A.M. (2014). The sense of recognition during retrieval failure: Implications for the nature of memory traces. In B.H. Ross’s Psychology of Learning and Motivation, Volume 60, pp. 77-112. Elsevier.
- Cleary, A.M. (2014). On the empirical study of déjà vu: Borrowing methodology from the study of the tip-of-the-tongue phenomenon. In B.L. Schwartz & A.S. Brown’s Tip-of-the-tongue States and Related Phenomena. . pp. 264-280. Cambridge University Press
- Cleary, A.M., Staley, S.R., & Klein, K.R. (2014). The effect of tip-of-the-tongue states on other cognitive judgments. In B.L. Schwartz & A.S. Brown’s Tip-of-the-tongue States and Related Phenomena. pp. 75-94. Cambridge University Press.
- Ryals, A.J., Cleary, A.M., & Seger, C.A. (2013). Recall versus familiarity when recall fails for words and scenes: The differential roles of the hippocampus, perirhinal cortex, and category-specific cortical regions. Brain Research, 1492, 72-91 .
- Ryals, A.J. & Cleary, A.M. (2012). The recognition without cued recall phenomenon: Support for a feature-matching theory over a partial recollection account. Journal of Memory and Language, 66, 747-762 .
- Cleary, A.M., Brown, A.S., Sawyer, B.D., Nomi, J.S., Ajoku, A.C., & Ryals, A.J. (2012). Familiarity from the configuration of objects in 3-dimensional space and its relation to déjà vu: A virtual reality investigation. Consciousness and Cognition, 21, 969-975 .
- Cleary, A. M., Ryals, A. J., & Nomi, J. N. (2009). Can déjà vu result from similarity to a prior experience? Support for the similarity hypothesis of déjà vu. Psychonomic Bulletin & Review, 16, 1082-1088 .
- Kostic, B. & Cleary, A. M. (2009). Song recognition without identification: When people cannot “name that tune” but can recognize it as familiar. Journal of Experimental Psychology: General, 138, 146-159 .
- Cleary, A.M. (2008). Recognition memory, familiarity, and déjà vu experiences. Current Directions in Psychological Science, 17, 353-357.
- Cleary, A. M. (2004). Orthography, phonology, and meaning: Word features that give rise to feelings of familiarity in recognition. Psychonomic Bulletin & Review, 11 , 446-451.
In the News:
BBC World Service – Deeply Human, Déjà vu
What’s Really Happening When You Experience Déjà Vu? | Discover Magazine
Déjà Vu For Your Ears | Wisconsin Public Radio (wpr.org)
Déjà vu links to the future – Psychonomic Society Featured Content
anne cleary | (ted.com)
BBC’s The Forum, 2015
Smithsonian Magazine, 2013
Scientific American, 2012

Mailing Address:
1876 Campus Delivery
Department of Psychology
Fort Collins, CO 80523-1876
Phone: 970-491-7701
FAX: 970-491-6363
Email: [email protected]
Anne_ClearyVitae (pdf)
- Apply to CSU
- Contact CSU
- Equal Opportunity
- Privacy Statement
THE STUDY OF HUMAN MEMORY
Boost your brain with mind lab pro.
Your brain is incredibly complex. Mind Lab Pro has 11 different nootropics all working together to increase your cognition and brainpower to help you live a better life.
If you need to perform at your best, need to focus, problem-solve or maintain a calm and clear mindset, you will get a huge benefit from taking Mind Lab Pro.
- Better focus
- Calm mindset
- 55+ memory and mood
- Performance focused athletes
- Student learning
The study of human memory stretches back at least 2,000 years to Aristotle ’s early attempts to understand memory in his treatise “On the Soul” . In this, he compared the human mind to a blank slate an d theorized that all humans are born free of any knowledge and are merely the sum of their experiences. Aristotle compared memory to making impressions in wax, sometimes referred to as the "storehouse metaphor" , a theory of memory which held sway for many centuries.
In antiquity, it was generally assumed that there were two sorts of memory: the “natural memory” (the inborn one that everyone uses every day) and the “artificial memory” (trained through learning and practice of a variety of mnemonic techniques , resulting in feats of memory that are quite extraordinary or impossible to carry out using the natural memory alone). Roman rhetoricians such as Cicero and Quintillian expanded on the art of memory or the method of loci (a method often first attributed to Simonides of Creos or the Pythagoreans), and their ideas were passed down to the medieval Scholastics and later scholars of the Renaissance like Matteo Ricci and Giordano Bruno .
The 18th Century English philosopher David Hartley was the first to hypothesize that memories were encoded through hidden motions in the nervous system, although his physical theory for the process was rudimentary at best. William James in America and Wilhelm Wundt in Germany, both considered among the founding fathers of modern psychology, both carried out some early basic research into how the human memory functions in the 1870s and 1880s (James hypothesized the idea of neural plasticity many years before it was demonstrated). In 1881, Théodule-Armand Ribot proposed what became known as Ribot's Law , which states that amnesia has a time-gradient in that recent memories are more likely to be lost than the more remote memories (although in practice this is actually not always the case).
However, it was not until the mid-1880s that the young German philosopher Herman Ebbinghaus developed the first scientific approach to studying memory. He did experiments using lists of nonsense syllables, and then associating them with meaningful words, and some of his findings from this work (such as the concepts of the learning curve and forgetting curve, and his classification of the three distinct types of memory : sensory , short-term and long-term ) remain relevant to this day.
The German evolutionary biologist Richard Semon first proposed in 1904 the idea that experience leaves a physical trace, which he called an engram, on specific webs of neurons in the brain. The British psychologist Sir Frederick Bartlett is considered one of the founding fathers of cognitive psychology, and his research in the 1930s into the recall of stories greatly influenced later ideas on how the brain stores memories.
With advances in technology in the 1940s, the field of neuropsychology emerged and with it a biological basis for theories of encoding. Karl Lashley devoted 25 years of his life to research on rats in mazes, in a systematic attempt to pinpoint where memory traces or engrams are formed in the brain, only to conclude in 1950 that memories are not localized to one part of the brain at all, but are widely distributed throughout the cortex, and that, if certain parts of the brain are damaged, other parts of the brain may take on the role of the damaged portion. The Canadian neurosurgeon Wilder Penfield ’s work on the stimulation of the brain with electrical probes in the 1940s and 1950s, initially in search of the causes of epilepsy, allowed him to create maps of the sensory and motor cortices of the brain that are still used today, practically unaltered. He was also able to summon up memories or flashbacks (some of which the patients had no conscious recollection of) by probing parts of the temporal lobe of the brain.
As early as 1949, another Canadian, Donald Hebb, intuited that “neurons that fire together, wire together”, implying that the encoding of memories occurred as connections between neurons were established through repeated use. This theoretical idea sometimes referred to as Hebb’s Rule was supported by the discovery of the mechanics of memory consolidation, long-term potentiation and neural plasticity in the 1970s, and remains the reigning theory today. Eric Kandel’s work on sea-slugs (whose brains are relatively simple and contain relatively large, and easily-observed individual neural cells) was particularly important in experimentally demonstrating Hebb’s Rule and identifying the molecular changes during learning, and the neurotransmitters involved.
As computer technology developed in the 1950s and 1960s, parallels between computer and brain processes became apparent, leading to advances in the understanding of the encoding, storage and retrieval processes of memory. The computer metaphor is, however, essentially just a more sophisticated version of the earlier storehouse view of memory, based on the rather simplistic and misleading assumption that memory is just a simple copy of the original experience.
The change in the overall study of memory during the 1950s and 1960s has come to be known as the “cognitive revolution”, and led to several new theories on how to view memory, and yielded influential books by George Miller, Eugene Galanter, Karl Pribram, George Sperling and Ulric Neisser. In 1956, George Miller produced his influential paper on short-term memory and his assessment that our short-term memory is limited to what he called “the magical number seven, plus or minus two”.
In 1968, Richard Atkinson and Richard Shiffrin first described their modal, or multi-store, a model of memory - consisting of a sensory memory , a short-term memory and a long-term memory - which became the most popular model for studying memory for many years. Fergus Craik and Robert Lockhart offered an alternative model, known as the levels-of-processing model, in 1972. In 1974, Alan Baddeley and Graham Hitch proposed their model of working memory, which consists of the central executive, visuospatial sketchpad and phonological loop as a method of encoding.
The 1970s also saw the early work of Elizabeth Loftus, who carried out her influential research on the misinformation effect, memory biases and the nature of false memories. The pioneering research on human memory by Endel Tulving from the 1970s onwards has likewise been highly influential. He was the first to propose two distinct kinds of long-term memory, episodic and semantic , in 1972 and he also devised the encoding specificity principle in 1983.
During the 1980s and 1990s, several formal models of memory were developed that can be run as computer simulations, including the Search of Associative Memory (SAM) model proposed by Jerome Raaijmaker and Richard Shiffrin in 1981, the Parallel Distributed Processing (PDP) model of James McClelland, David Rumelhart and Geoffrey Hinton's in 1986, and various versions of the Adaptive Control of Thought (ACT) model developed by John Anderson in 1993.
Nowadays, the study of human memory is considered part of the disciplines of cognitive psychology and neuroscience , and the interdisciplinary link between the two which is known as cognitive neuroscience . You can visit https://www.advancedwriters.com/custom-research-paper/ if you need research paper help from experts.
- International edition
- Australia edition
- Europe edition

Write down your thoughts and shred them to relieve anger, researchers say
Writing negative reactions on paper and shredding it or scrunching and throwing in the bin eliminates angry feelings, study finds
Since time immemorial humans have tried to devise anger management techniques.
In ancient Rome, the Stoic philosopher Seneca believed “my anger is likely to do me more harm than your wrong” and offered avoidance tips in his AD45 work De Ira (On Anger).
More modern methods include a workout on the gym punchbag or exercise bike. But the humble paper shredder may be a more effective – and accessible – way to decompress, according to research.
A study in Japan has found that writing down your reaction to a negative incident on a piece of paper and then shredding it, or scrunching it into a ball and throwing it in the bin, gets rid of anger.
“We expected that our method would suppress anger to some extent,” said Nobuyuki Kawai, lead researcher of the study at Nagoya University. “However, we were amazed that anger was eliminated almost entirely.”
The study, published in Scientific Reports on Nature , builds on research on the association between the written word and anger reduction as well as studies showing how interactions with physical objects can control a person’s mood. For instance, those wanting revenge on an ex-partner may burn letters or destroy gifts.
Researchers believe the shredder results may be related to the phenomenon of “backward magical contagion”, which is the belief that actions taken on an object associated with a person can affect the individuals themselves. In this case, getting rid of the negative physical entity, the piece of paper, causes the original emotion to also disappear.
This is a reversal of “magical contagion” or “celebrity contagion” – the belief that the “essence” of an individual can be transferred through their physical possessions.
Fifty student participants were asked to write brief opinions about an important social problem, such as whether smoking in public should be outlawed. Evaluators then deliberately scored the papers low on intelligence, interest, friendliness, logic, and rationality. For good measure, evaluators added insulting comments such as: “I cannot believe an educated person would think like this. I hope this person learns something while at the university.”
The wound-up participants then wrote down their angry thoughts on the negative feedback on a piece of paper. One group was told to either roll up the paper and throw it in a bin or keep it in a file on their desk. A second group was told to shred the paper, or put it in a plastic box.
Anger levels of the individuals who discarded their paper in the bin or shredded it returned to their initial state, while those who retained a hard copy of the paper experienced only a small decrease in their overall anger.
Researchers concluded that “the meaning (interpretation) of disposal plays a critical role” in reducing anger.
“This technique could be applied in the moment by writing down the source of anger as if taking a memo and then throwing it away,” said Kawai.
Along with its practical benefits, this discovery may shed light on the origins of the Japanese cultural tradition known as hakidashisara ( hakidashi sara refers to a dish or plate) at the Hiyoshi shrine in Kiyosu, just outside Nagoya. Hakidashisara is an annual festival where people smash small discs representing things that make them angry. The study’s findings may explain the feeling of relief that participants report after leaving the festival, the paper concluded.

Fridge magnets can be cool aid to holiday memory recall, study finds

Want to skip that Christmas party? The host probably won’t mind, study shows

‘Succession syndrome’ prevalent among wealthy households, psychiatrists warn

Drugs and alcohol do not make you more creative, research finds

Why being rude to the waiter (or other staff) is the worst strategy

Authors of original dating profiles rated more attractive, research finds

I fear my children are overexposed to technology. Experts say I’m right to worry

I used to be ashamed of being a fangirl. Now I see how joyous and creative it was

Autistic scholar Temple Grandin: ‘The education system is screening out visual thinkers’

Human neurons transplanted into rats to help study brain disorders
Most viewed.

Study Maps The Odd Structural Similarities Between The Human Brain And The Universe
A lthough they seem vastly different, the human brain and the universe actually have a surprising amount in common. Scientists have demonstrated this before, in an image that occasionally makes its rounds on the internet. The photo shows a human neuron and a simulated galaxy cluster. Side-by-side, the two look shockingly alike [ 1 ].
Two scientists from Italy took that comparison one step further. In their new pilot study, they compare the neuronal network in the human brain with the cosmic network of the galaxies in the universe. All told, these are two of the most complex systems in nature.
The Human Brain and the Universe
The two scientists responsible for the study are astrophysicist Franco Vazza of the University of Bologna, and Alberto Feletti of the University of Verona, both in Italy. Their purpose was to find out if the similarities between the human brain and the universe were more than just by the way they look.
With a hundred billion neurons, and a hundred trillion connections, the human brain is one of the most complex objects in our universe. That made Vazza and Feletti wonder- is the human brain more complicated than the galaxies in our universe?
"Galaxies can group into enormous structures (called clusters, superclusters, and filaments) that stretch for hundreds of millions of light-years," they explained in Nautilus in 2017. "The boundary between these structures and neighboring stretches of empty space called cosmic voids can be extremely complex." [ 2 ]
They continued to explain that considering the number of bits of information it takes to describe it, the void-filament boundary is one of the most complex volumes in the universe. So which is more complex- the brain or the universe?
Read: Scientists spot a ‘kilonova' flash so bright they can barely explain it
How Did They Compare the Human Brain and the Universe?
To answer their question, the team used quantitative analysis. They based their analysis on images from slices of the human cerebellum and cortex at different magnifications. They then compared these to simulations of the cosmic web.
Specifically, they were looking for matter density fluctuations between brains and the cosmic web. Given the overwhelmingly large size difference between the brain and the universe, these relative fluctuations occur on vastly different scales. Despite this, the similarities between the two were striking [ 3 ].
What Did They Find?
"We calculated the spectral density of both systems. This is a technique often employed in cosmology for studying the spatial distribution of galaxies" , explained Vazza. "Our analysis showed that the distribution of the fluctuation within the cerebellum neuronal network on a scale from 1 micrometer to 0.1 millimeters follows the same progression of the distribution of matter in the cosmic web but, of course, on a larger scale that goes from 5 million to 500 million light-years". [ 4 ]
So our brains and the universe have a similar distribution of matter, but what else do they have in common? The physical processes that create the structure of the human brain and that of the universe are extremely different. The researchers said, however, that they can result in similar levels of complexity and self-organization.
Press These Points: 18 Self-Help Tips for Sciatica That Truly Work
The list of commonalities between the human brain and the universe, then, is surprisingly long:
- The human cerebellum has around 69 billion neurons. The cosmic galaxy contains over 100 billion galaxies. In the grand scheme of things, those numbers aren't wildly different.
- Distinct networks make up both systems, each containing nodes. In the brain, we call those nodes neurons, in the universe, we call them galaxies. In both cases, filaments connect these nodes.
- The radius of both neurons and galaxies are only a fraction of the length of the filaments. The flow of energy and information between the nodes is only about 25 percent of the mass and energy content of each system.
- The brain is about 77 percent water, the universe is about 72 percent dark energy. While both of these materials appear to be passive, they play an indirect role in their internal structures [ 5 ].
Read: For the First Time Ever, Scientists Caught Time Crystals Interacting
The Brain and the Universe: Connections and Memory
The team already knew many of these similarities before their research. So what else did they find?
They learned that the cosmic web had an average of 3.1 to 4.8 connections per node. The human cortex was not that dissimilar. It had an average of 4.6 to 5.4 connections per node. Both systems tended to cluster their connections around central nodes, and both had a similar information capacity [5].
Additionally, a 2015 study calculated that the memory of the human brain is about 2.5 petabytes [ 6 ]. Vazza conducted another study in 2019 which found that the memory capacity needed to store the complexity of the universe is approximately 4.3 petabytes [ 7 ].
The Brain and the Universe: More Similarities than Differences?
The universe, of course, does not have a sentient brain the way humans do. The similarities in memory capacity between the human brain and the universe, however, may indicate that the laws that govern their growth and structure could be the same [2].
"Once again, structural parameters have identified unexpected agreement levels," said Feletti. "Probably, the connectivity within the two networks evolves following similar physical principles, despite the striking and obvious difference between the physical powers regulating galaxies and neurons." [5]
This study by Vazza and Feletti, along with other similar studies, are very important. The hope is that they will help scientists better understand the laws that govern both our brains and the universe.
These two vastly different networks, says Feletti, show more similarities than those shared between a cosmic web and a galaxy, or a neuronal network and the inside of a neuronal body. With the knowledge, the more we learn about one, the more we may be able to learn about the other.
Keep Reading: Mystery Meteorite Contains "Impossible to Naturally Exist" Crystal
- https://sprott.physics.wisc.edu/pickover/pc/brain-universe.html
- https://nautil.us/issue/50/emergence/the-strange-similarity-of-neuron-and-galaxy-networks
- https://www.frontiersin.org/articles/10.3389/fphy.2020.525731/full#B6
- https://www.eurekalert.org/pub_releases/2020-11/udb-dth111620.php
- https://www.sciencealert.com/wildly-fun-new-paper-compares-the-human-brain-to-the-structure-of-the-universe
- https://elifesciences.org/articles/10778
- https://academic.oup.com/mnras/article-abstract/491/4/5447/5644441?redirectedFrom=fulltext
The post Study Maps The Odd Structural Similarities Between The Human Brain And The Universe appeared first on The Hearty Soul .
17+ Hygiene Habits That Could Be Harming You Daily

AI Index Report
The AI Index Report tracks, collates, distills, and visualizes data related to artificial intelligence. Our mission is to provide unbiased, rigorously vetted, broadly sourced data in order for policymakers, researchers, executives, journalists, and the general public to develop a more thorough and nuanced understanding of the complex field of AI. The report aims to be the world’s most credible and authoritative source for data and insights about AI.
Subscribe to receive the 2024 report in your inbox!

Coming Soon: 2024 AI Index Report!
The 2024 AI Index Report will be out April 15! Sign up for our mailing list to receive it in your inbox.
Steering Committee Co-Directors

Ray Perrault
Steering committee members.

Erik Brynjolfsson

John Etchemendy

Katrina Ligett

Terah Lyons

James Manyika

Juan Carlos Niebles

Vanessa Parli

Yoav Shoham

Russell Wald
Staff members.

Loredana Fattorini

Nestor Maslej
Letter from the co-directors.
AI has moved into its era of deployment; throughout 2022 and the beginning of 2023, new large-scale AI models have been released every month. These models, such as ChatGPT, Stable Diffusion, Whisper, and DALL-E 2, are capable of an increasingly broad range of tasks, from text manipulation and analysis, to image generation, to unprecedentedly good speech recognition. These systems demonstrate capabilities in question answering, and the generation of text, image, and code unimagined a decade ago, and they outperform the state of the art on many benchmarks, old and new. However, they are prone to hallucination, routinely biased, and can be tricked into serving nefarious aims, highlighting the complicated ethical challenges associated with their deployment.
Although 2022 was the first year in a decade where private AI investment decreased, AI is still a topic of great interest to policymakers, industry leaders, researchers, and the public. Policymakers are talking about AI more than ever before. Industry leaders that have integrated AI into their businesses are seeing tangible cost and revenue benefits. The number of AI publications and collaborations continues to increase. And the public is forming sharper opinions about AI and which elements they like or dislike.
AI will continue to improve and, as such, become a greater part of all our lives. Given the increased presence of this technology and its potential for massive disruption, we should all begin thinking more critically about how exactly we want AI to be developed and deployed. We should also ask questions about who is deploying it—as our analysis shows, AI is increasingly defined by the actions of a small set of private sector actors, rather than a broader range of societal actors. This year’s AI Index paints a picture of where we are so far with AI, in order to highlight what might await us in the future.
- Jack Clark and Ray Perrault
Our Supporting Partners

Analytics & Research Partners

Stay up to date on the AI Index by subscribing to the Stanford HAI newsletter.
share this!
April 9, 2024
This article has been reviewed according to Science X's editorial process and policies . Editors have highlighted the following attributes while ensuring the content's credibility:
fact-checked
peer-reviewed publication
trusted source
Can large language models replace human participants in some future market research?
by Institute for Operations Research and the Management Sciences

Do market researchers still need to conduct original research using human participants in their work? Not always, according to a new study. The study found that thanks to the increasing sophistication of large language models (LLMs), human participants can be substituted with LLMs and still generate similar outputs as those generated from human surveys.
The study is published in Marketing Science . The article, "Determining the Validity of Large Language Models for Automated Perceptual Analysis," is authored by Peiyao Li and Zsolt Katona, both of the University of California, Berkeley; Noah Castelo of the University of Alberta and Miklos Sarvary of Columbia University.
According to the research, agreement rates between human- and LLM-generated data sets reached 75%–85%.
"LLMs can be used to generate text when given a prompt on certain generative Artificial Intelligence (AI) platforms," says Li. "Our research focused on perceptual analysis and the use of automated market research for certain product categories."
To conduct their research, the study authors used LLMs to tap data that is broadly available on the internet. They developed a new methodology and workflow that allows market researchers to rely only on an LLM to conduct market research. As a result, they demonstrated that LLM-powered market research can produce meaningful results and even replicate human results.
"It is important to note that with LLMs, while market researchers may not require interviews with human research subjects, the ultimate data does originate from human beings, using available data," says Katona. "LLMs have been engineered to accurately replicate human responses based on machine learning of actual human perceptions, attitudes, and preferences."
Castelo added, "The core LLM takes a prompt as an input and generates a continuation of text as output. With proper prompting, the LLM can then generate comparisons and assessments of various brands or products in a given category and produce results that are, at the moment, 75%–85% in agreement with research featuring human participants ."
The researchers believe that for some product and brand categories, their new method of fully or partially automating market research will increase the efficiency of market research by speeding up the process and potentially reducing cost. At the same time, they caution that fully automated market research without human input may not be accurate for all product categories.
"While we are very excited about the possibilities we've seen through our research, we recognize that this is just the beginning and going forward, LLM-based market research will be able to answer more nuanced questions as the market research field begins to tap and develop its potential," says Sarvary.
Explore further
Feedback to editors

Metasurface antenna could enable future 6G communications networks
41 minutes ago

Making cement is very damaging for the climate. One solution is opening in California
8 hours ago

New computer vision tool can count damaged buildings in crisis zones and accurately estimate bird flock sizes
20 hours ago

Engineers recreate Star Trek's Holodeck using ChatGPT and video game assets
21 hours ago

Engineers quicken the response time for robots to react to human conversation
Apr 11, 2024

Researchers show electrical pulses can control thermal resistance in devices

New open-source generative machine learning model simulates future energy-climate impacts

Adding a telescopic leg beneath a quadcopter to create a hopping drone

Tiny AI-trained robots demonstrate remarkable soccer skills

Discovery brings all-solid-state sodium batteries closer to practical use
Related stories.

Large language models in health: Useful, but not a miracle cure
Apr 3, 2024

Who wrote this? Engineers discover novel method to identify AI-generated text
Mar 20, 2024

Large language models pose risk to science with false answers, says study
Nov 20, 2023

Trust your doctor: Study shows human medical professionals are more reliable than artificial intelligence tools
Apr 2, 2024

Study argues that large language models can reveal breakthroughs in neuroscience that humans alone cannot
Feb 9, 2024

AI can 'lie and BS' like its maker, but still not intelligent like humans, argues researcher
Nov 21, 2023
Recommended for you

AI-powered 'sonar' on smartglasses tracks gaze, facial expressions
Apr 10, 2024
Let us know if there is a problem with our content
Use this form if you have come across a typo, inaccuracy or would like to send an edit request for the content on this page. For general inquiries, please use our contact form . For general feedback, use the public comments section below (please adhere to guidelines ).
Please select the most appropriate category to facilitate processing of your request
Thank you for taking time to provide your feedback to the editors.
Your feedback is important to us. However, we do not guarantee individual replies due to the high volume of messages.
E-mail the story
Your email address is used only to let the recipient know who sent the email. Neither your address nor the recipient's address will be used for any other purpose. The information you enter will appear in your e-mail message and is not retained by Tech Xplore in any form.
Your Privacy
This site uses cookies to assist with navigation, analyse your use of our services, collect data for ads personalisation and provide content from third parties. By using our site, you acknowledge that you have read and understand our Privacy Policy and Terms of Use .
E-mail newsletter

Date & time
Street 3, The Street Theatre
Event description
Memory in the City
Cities are not only brick and mortar. In cities, cultures converge, interact, flourish, and sometimes clash violently. Cities hold the key to restoring and making whole the fractures and silences of our collective memory and identity, shaping how we see ourselves and, with it, our home and the world around us.
'Memory in the City’ celebrates the power of creative and historical writing and art to reimagine cities as places of memory, solidarities, narratives, and new perspectives.
Through an imaginative and interactive reading of André Dao's Anam, the author, and ANU scholars will take us on a journey through different urban landscapes – Melbourne, Hanoi, Paris, and Cambridge, to explore the long-lasting impact of migration on the cultural tapestry of our cities and the potential for new pasts and futures.
Moderator, Ann-Sophie Levidis , Lecturer in French Studies, ANU Andre Dao , Refugee Advocate and Novelist Leslie Barnes , Associate Professor in French Studies, ANU Desmond Manderson , Professor of Law, Director of the Centre for Law Arts and Humanities, ANU Sue Thompson , Associate Professor of Southeast Asian Studies, ANU
Wellspring: Enquiry and Exchange Bringing campus to the community, Wellspring is an exciting series of immersive events that explores new depths of cultural enquiry and exchange. A collaboration between The Street Theatre and the ANU School of Literature, Languages and Linguistics, Wellspring invites Canberra’s curious to experience the inexhaustible source of human creativity and communication. Get ready for big bold ideas inviting curiosity and conversation.
More information
- Skip to main content
- Keyboard shortcuts for audio player

- LISTEN & FOLLOW
- Apple Podcasts
- Google Podcasts
- Amazon Music
- Amazon Alexa
Your support helps make our show possible and unlocks access to our sponsor-free feed.
What to know about the new EPA rule limiting 'forever chemicals' in tap water

Berly McCoy

Regina G. Barber

Rebecca Ramirez

Following a new EPA rule, public water systems will have five years to address instances where there is too much PFAS in tap water – three years to sample their systems and establish the existing levels of PFAS, and an additional two years to install water treatment technologies if their levels are too high. Justin Sullivan/Getty Images hide caption
Following a new EPA rule, public water systems will have five years to address instances where there is too much PFAS in tap water – three years to sample their systems and establish the existing levels of PFAS, and an additional two years to install water treatment technologies if their levels are too high.
Wednesday the Environmental Protection Agency announced new drinking water standards to limit people's exposure to some PFAS chemicals.
For decades, PFAS have been used to waterproof and stain-proof a variety of consumer products. These "forever chemicals" in a host of products — everything from raincoats and the Teflon of nonstick pans to makeup to furniture and firefighting foam. Because PFAS take a very long time to break down, they can accumulate in humans and the environment.

Shots - Health News
Epa puts limits on 'forever chemicals' in drinking water.

'Forever chemicals' could be in nearly half of U.S. tap water, a federal study finds
Now, a growing body of research is linking them to human health problems like serious illness, some cancers, lower fertility and liver damage.
Science correspondent Pien Huang joins the show today to talk through this new EPA rule — what the threshold for safe levels of PFAS in tap water is, why the rule is happening now and how the federal standards will be implemented.
Listen to Short Wave on Spotify , Apple Podcasts and Google Podcasts .
Listen to every episode of Short Wave sponsor-free and support our work at NPR by signing up for Short Wave+ at plus.npr.org/shortwave .
This episode was produced by Berly McCoy. It was edited by Rebecca Ramirez and Scott Hensley. Rebecca, Berly and Pien Huang checked the facts.
- forever chemicals
- drinking water
- Environmental Protection Agency
IMAGES
VIDEO
COMMENTS
However, further research is needed to study the molecular and structural changes brought on by implicit memory (Gallistel and Balsam, 2014). The contribution of non-human animal studies toward our understanding of memory processes cannot be understated; hence recognizing their value is vital for moving forward.
Study sheds light on how neurons form long-term memories. On a late summer day in 1953, a young man who would soon be known as patient H.M. underwent experimental surgery. In an attempt to treat his debilitating seizures, a surgeon removed portions of his brain, including part of a structure called the hippocampus. The seizures stopped.
The goal of the present ar cle is to present and discuss a. series of open ques ons relat ed to major topics on human memory research that can be addressed by future research. The. topics covered ...
The Neuropsychology of Human Learning and Memory Felicia B. Gershberg Memory Disorders Research Center, Boston University School of Medicine and Department of Veterans Affairs Medical Center, Boston, Massachusetts 02130 Arthur P. Shimamura Department of Psychology, University of California, Berkeley, California 94720 I. INTRODUCTION Neuropsychological descriptions of memory disorders date back ...
The history of human memory research might have taken a different turn if neurosurgeon William Scoville had not removed both hippocampi in one patient to treat his seizure disorder. ... There are numerous methods now available to study human memory, which differ in several important ways. While some methods are more suitable to localize ...
In this special issue of Nature Neuroscience, we feature an assortment of reviews and perspectives that explore the topic of learning and memory. Learning new information and skills, storing this ...
Human memory: An introduction to research, data, and theory. 2d ed. Belmont, CA: Wadsworth. This textbook is aimed at advanced undergraduate students and provides a comprehensive foundation for the study of human memory, including an introduction to mathematical and computational models of human memory.
Abstract. Due to the advent of neuropsychology, it has become clear that there is a multiplicity of memory systems or, at the very least, of dissociably different modes of processing memory in the brain. As the Oxford Handbook of Memory demonstrates, the frontier of memory research has been enriched by breakthroughs of the last decades, with ...
The study of memory incorporates research methodologies from neuropsychology, human development and animal testing using a wide range of species. The complex phenomenon of memory is explored by combining evidence from many areas of research. New technologies, experimental methods and animal experimentation have led to an increased understanding of the workings of memory.
Abstract. Understanding the complex relationship between sleep and memory consolidation is a major challenge in cognitive neuroscience and psychology. Many studies suggest that sleep triggers off ...
Memory and cognition: shifting approaches to how we do science. On 1 January 2020, I took on the role as Editor-in-Chief of Memory and Cognition. My vision has been and continues to be to create a venue for research that broadens our understanding of human cognition. I seek to do this by encouraging more research that examines cognition in ...
1. Recognition memory: Considering Tulving's contributions and some new findings. Endel Tulving's contributions to the psychology and cognitive neuroscience of memory are vast. The task we have undertaken here is to focus on a few of his contributions to the study of recognition memory and its relation to recall.
Memory publishes high quality papers in all areas of memory research, ... provides a unique venue for memory researchers to communicate their findings and ideas both to peers within their own research tradition in the study of memory, and also to the wider range of research communities with direct interest in human memory. Peer Review: All ...
In this lab, we study human memory. Though we study a broad range of issues related to human memory processes and performance, the primary research interest in this lab is in people's ability to show memory when they experience retrieval failure. For example, people sometimes fail to recall the information that they are searching for in memory.
The advent of virtual environments (VEs) has given us a new kind of tool for the studies of cognitive issues. In order to develop a satisfactory research methodology, we need to know more about the implication of human factors interacting with virtual reality. Rules of both physical motoric interfaces and visual displays on cognitive function as memory are not completely clear. In this study ...
The pioneering research on human memory by Endel Tulving from the 1970s onwards has likewise been highly influential. He was the first to propose two distinct kinds of long-term memory, episodic and semantic, in 1972 and he also devised the encoding specificity principle in 1983. During the 1980s and 1990s, several formal models of memory were
Memory reactivation during sleep is thought to facilitate memory consolidation. Most sleep reactivation research has examined how reactivation of specific facts, objects, and associations benefits their overall retention. However, our memories are not unitary, and not all features of a memory persist in tandem over time. Instead, our memories are transformed, with some features strengthened ...
The prefrontal-cortex basal ganglia working memory (PBWM) model (Hazy et al., 2007; O'Reilly & Frank, 2006) proposes that working memory representations are updated via a striatal gating mechanism but lacks conclusive empirical support for the postulated subcortical involvement. A growing body of research suggests that dopamine is also involved in working memory updating (Braver & Cohen, 2000 ...
Introduction: White matter hyperintensities of presumed vascular origin (WMH) are associated with cognitive impairment and are a key imaging marker in evaluating brain health. However, WMH volume alone does not fully account for the extent of cognitive deficits and the mechanisms linking WMH to these deficits remain unclear. Lesion network mapping (LNM) enables to infer if brain networks are ...
A study in Japan has found that writing down your reaction to a negative incident on a piece of paper and then shredding it, or scrunching it into a ball and throwing it in the bin, gets rid of anger.
The Human Brain and the Universe. The two scientists responsible for the study are astrophysicist Franco Vazza of the University of Bologna, and Alberto Feletti of the University of Verona, both ...
The AI Index Report tracks, collates, distills, and visualizes data related to artificial intelligence. Our mission is to provide unbiased, rigorously vetted, broadly sourced data in order for policymakers, researchers, executives, journalists, and the general public to develop a more thorough and nuanced understanding of the complex field of AI.
Castelo added, "The core LLM takes a prompt as an input and generates a continuation of text as output. With proper prompting, the LLM can then generate comparisons and assessments of various brands or products in a given category and produce results that are, at the moment, 75%-85% in agreement with research featuring human participants.". The researchers believe that for some product and ...
Memory in the CityCities are not only brick and mortar. In cities, cultures converge, interact, flourish, and sometimes clash violently. Cities hold the key to restoring and making whole the fractures and silences of our collective memory and identity, shaping how we see ourselves and, with it, our home and the world around us.'Memory in the City' celebrates the power of creative and ...
'Forever chemicals' could be in nearly half of U.S. tap water, a federal study finds. Now, a growing body of research is linking them to human health problems like serious illness, some cancers ...