
Nutrition in Bacteria
Nutrition in bacteria is primarily categorized into two major groups, namely autotrophic and heterotrophic. Classification of bacteria in terms of nutritional requirement typically depends on the energy sources , whether in the form of light or chemical energy. Some bacteria obtain energy directly from the sunlight. However, a few bacteria get chemical energy by utilizing inorganic and organic carbon compounds as their sole energy source.
Autotrophic bacteria are photosynthetic organisms that can prepare food for themselves in the presence of sunlight and bacteriochlorophyll pigments. Oppositely, heterotrophic bacteria represent a group of organisms that depend on another host for food and survival.
Therefore, bacteria’s heterotrophic mode of nutrition is classified further into saprophytic, symbiotic and parasitic modes of nutrition based on the feeding mechanisms and the host type. This post describes the definition of nutrition and nutritional types of bacteria with examples.
Content: Nutrition in Bacteria
Definition of nutrition.
- Nutrient Sources
- Energy Sources
- Nutritional Types
Nutrition is a process in which living organisms acquire energy and nutrition through their surroundings to carry out various cellular processes. All living organisms have different nutritional requirements for growth and development. Bacteria are prokaryotic organisms that need energy and nutrients to undergo activities like growth, reproduction, and metabolism.
Nutrient Sources in Bacteria
Bacteria necessarily require macronutrients (such as carbon, nitrogen, hydrogen, oxygen, phosphorus, and sulfur) to build important biomolecules (proteins and nucleic acids) that constitute most cellular components.
- Carbon is the essential macronutrient that constitutes a significant portion of the cell. It builds biomolecules like carbohydrates, proteins, nucleic acids, lipids, etc.
- Nitrogen is necessary to build proteins and nucleic acids. Bacteria may utilize atmospheric nitrogen or other inorganic forms.
- Hydrogen and oxygen constitute many organic compounds, and they are part of water.
- Phosphorus is necessary to synthesize nucleotides and phospholipids.
- Sulfur is a part of amino acids (like cysteine, methionine).
- Besides all of the above, potassium, magnesium, calcium and sodium are essential to maintain the bacterial cell structure and function.
Bacteria need micronutrients (like iron, boron, chromium, and manganese) in trace or minute amounts to perform electron transport chain reactions and other processes.
- Bacteria need iron for the functioning of cytochromes and to undergo electron transport reactions.
- Some bacteria utilize elements like boron , chromium and manganese as they function as primary cofactors.
Energy Sources in Bacteria
They obtain energy from direct light and the oxidation-reduction reactions of different chemical compounds present in our environment. Depending upon the energy sources, the nutrition in bacteria is mainly autotrophic and heterotrophic. Phototrophic bacteria possess bacteriochlorophyll pigments to synthesize food, while heterotrophic bacteria rely on another host for food and energy.
Nutritional Types of Bacteria
Depending upon the energy, electron and carbon sources, bacteria can be classified into different groups, as mentioned in the diagram below. Phototrophs and chemotrophs are the two groups of bacteria that obtain energy through direct sunlight and chemicals.
Then, lithotrophs and organotrophs represent a bacterial group that use reduced inorganic carbon and organic compounds as electron donors , respectively. At last, bacteria utilizing carbon sources are grouped into autotrophs and heterotrophs.

Based on Energy Source
- Chemotrophs includes organisms that rely on chemical energy obtained through the oxidation of chemical compounds. Such organisms use a chemosynthesis mechanism where they use the energy released by the oxidation of inorganic molecules to make organic substances. Based on the cellular carbon source , chemoautotrophs and chemoheterotrophs are the two common types. Conversely, chemoorganotrophs and chemolithotrophs are the two groups of chemotrophs based on the origin of a hydrogen donor .
- Phototrophs include organisms that utilize light energy or capture energy photons. Such organisms use a photosynthesis mechanism where they use light energy to make organic molecules. Based on the cellular carbon source , photoautotrophs and photoheterotrophs are the two common types.
Based on Electron Source
- Chemo-lithotrophic bacteria gain energy from the reduced inorganic compounds. For example, Nitrosomonas use NH 3 as a source of an electron.
- Photo-lithotrophic bacteria gain energy from light and use reduced inorganic compounds. For example, Chromatium okeinii uses H 2 S as an electron source.
- Photo-organotrophic bacteria capture light energy and utilize organic compounds. For example, Rhodospirillum uses succinate as a source of an electron.
- Chemo-organotrophic bacteria solely obtain energy from organic compounds. For example, Pseudomonas pseudoflora use glucose and amino acids as a source of an electron.
Based on Carbon Source
They typically make food from inorganic sources (H 2 O, CO 2 , H 2 S salts). The autotrophic bacteria are of two types:
- Photoautotrophs capture the photons of sunlight through bacteriochlorophylls and convert them into chemical energy. They generally reduce CO 2 to carbohydrates. Inorganic substances serve as hydrogen donors.
- Unlike photoautotrophs, chemoautotrophs do not require light and chlorophyll pigments. These bacteria undergo oxidation-reduction reactions of certain inorganic substances. As a result of this, the energy is released or the reaction is exothermic. Chemoautotrophic bacteria derive this energy to carry out the synthetic processes of the cell.
Heterotrophs
They obtain their-ready made food from organic substances, living or dead. These are of two types:
- Photoheterotrophs harness light energy but do not utilize CO 2 as the sole carbon source. They fulfil carbon and electron requirements through organic compounds. Such bacteria possess bacteriochlorophyll pigment.
- Saprophytic bacteria obtain nutrition from dead organic matter such as decomposed leaves and vegetables. These bacteria secrete enzymes to degrade complex food, such as carbohydrates and protein, into simple absorbable compounds. Examples are Bacillus mycoides , B. ramosus , Acetobacter etc.
- Parasitic bacteria obtain their nutrition from the living host tissues. They may be harmless or deleterious that may cause severe illnesses and diseases. Examples are Rickettsia , Chlamydia , Bacillus typhosus , Vibrio cholerae , Pseudomonas citri etc.
- Symbiotic bacteria are closely associated with some organisms called symbionts. They are beneficial to organisms. The common examples are the nitrogen-fixing bacteria (like Rhizobium , Clostridium etc.), those living inside the roots of leguminous plants. Nitrifiers perform a primary role in fixing free atmospheric nitrogen into nitrogenous compounds, which plants utilize. In return, the plant harbours the bacteria by giving essential nutrients and a favourable environment for growth.
Therefore, we can conclude that the nutritional types of bacteria rely upon different energy sources like light energy and chemical energy to undergo various cellular processes.
Related Topics:
- Electrophoresis
- Difference Between Thymine and Uracil
- Hershey and Chase Experiment
- Indole Test
- Reaction Centre
1 thought on “Nutrition in Bacteria”
Nutrition is the study of how food and nutrients impact our bodies and health. A well-balanced diet is essential to maintain good health, prevent chronic diseases, and promote longevity. There are different types of nutrition, including autotrophic, heterotrophic, mixotrophic, and malnutrition.
Leave a Comment Cancel Reply
Your email address will not be published. Required fields are marked *
Start typing and press enter to search

Username or Email Address
Remember Me Forgot Password?
Get New Password
Classification of Bacteria on the Basis of Nutrition with examples
- By Mubashir Iqbal
- On June 15, 2023
- In Basics of Microbiology
Table of Contents
Introduction
An essential component of understanding the large diversity of microorganisms that live in our world is bacterial classification. By classifying bacteria according to their nutritional requirements, we learn a great deal about their metabolic capacities and ecological functions. This article attempts to investigate how bacteria are categorized based on nutrition, illuminating the numerous varieties and their importance in varied contexts.
Autotrophic Bacteria
A group of microorganisms known as autotrophic bacteria is capable of synthesizing their own organic compounds from inorganic materials. They differ from heterotrophic bacteria, which depend on external organic resources for nutrition, by this process, known as autotrophy . The distinctive feature of autotrophic bacteria is the use of inorganic carbon as the main source of carbon for growth.
Photosynthetic Autotrophic Bacteria
Oxygenic photosynthetic bacteria.
- Cyanobacteria, a group of autotrophic microorganisms that produce oxygen through the process of photosynthesis.
- these bacteria use the light energy to transform carbon dioxide into organic chemicals.
- Chlorophyll pigments are used in oxygenic photosynthesis to capture light energy and start the electron transport chain, which results in the generation of oxygen as a byproduct.
- A variety of ecosystems, including freshwater, marine, and even symbiotic interactions with plants, are habitat to cyanobacteria.
- Examples: Cyanobacteria, Prochlorococcus
Anoxygenic Photosynthetic Bacteria
- The type of autotrophic microorganisms that can use light energy to synthesize organic molecules is called anoxygenic photosynthetic bacteria .
- They do not produce oxygen as a byproduct, in contrast to oxygenic photosynthetic bacteria. The pigments bacteriochlorophylls, bacterioviridins , or bacteriorhodopsins are just a few of the pigments used by anoxygenic photosynthetic bacteria to absorb light energy and fuel their metabolic processes.
- These bacteria may survive in a variety of environments, including the anaerobic habitats present in aquatic sediments, hot springs, and even animal digestive tracts.
- Examples: Purple sulfur bacteria, Green non-sulfur bacteria
Chemolithotrophic Autotrophic Bacteria
A group of microorganisms known as chemolithotrophic autotrophic bacteria or chemolithoautotrophic bacteria obtains their energy from the oxidation of inorganic substances. Instead of using light or organic molecules as an energy source, these bacteria use a variety of inorganic substrates. They can survive and even thrive in conditions without organic matter, like sulfur-rich hot springs or deep-sea hydrothermal vents.
Sulfur-Oxidizing Bacteria
- Sulfur-oxidizing bacteria are chemolithotrophic autotrophs that oxidize inorganic sulfur compounds such as thiosulfate (S2O3-), elemental sulfur ( S0), and hydrogen sulfide (H2S).
- They carry out these oxidation reactions using specialized enzymes like sulfur oxygenases or thiosulfate reductases.
- Sulfur-oxidizing bacteria contribute to the transformation of sulfide minerals into sulfate and are essential for sulfur cycling in both marine and terrestrial environments. They play a vital part in biogeochemical cycles.
- Examples: Thiobacillus, Beggiatoa
Iron Oxidizing Bacteria
- Iron-oxidizing bacteria are chemolithotrophic autotrophs that get their energy from oxidizing ferrous iron (Fe2+) .
- These bacteria have special enzymes called rusticyanins or proteins that resemble rusticyanins, which promote the oxidation of iron.
- Acidic mine drainage and iron-rich soils are two examples of habitats where iron-oxidizing bacteria are frequently found.
- They have an impact on the creation of iron-rich deposits and are crucial to the iron cycle.
- Examples: Gallionella, Leptothrix
Nitrogen fixing Bacteria
- Nitrogen-fixing bacteria are chemolithotrophic autotrophs with the ability to fix nitrogen in the atmosphere (N2) to produce ammonia (NH3).
- This process is critical to the nitrogen cycle because it allows nitrogen to be incorporated into organic molecules.
- In symbiotic partnerships with plants like legumes, nitrogen-fixing bacteria live inside specialized structures called nodules and supply the plant with an essential source of nitrogen for growth.
- Examples: Rhizobium, Azotobacter
Heterotrophic bacteria
Heterotrophic bacteria are a wide group of microorganisms that get their carbon and energy from organic sources. They cannot create their own organic compounds, unlike autotrophic bacteria, so they must obtain nutrients from external organic materials. They then break these complex organic molecules down into simpler compounds to produce energy and create cellular components.
Aerobic heterotrophic bacteria
Saprophytic bacteria.
- Aerobic heterotrophic bacteria that specialize in digesting decaying organic substances are called saprophytic bacteria.
- They are essential for recycling of organic materials and the cycling of nutrients in ecosystems.
- Extracellular enzymes secreted by saprophytic bacteria help break down complex organic materials into simpler molecules that may be easily absorbed and used by the bacterium for growth and energy production.
- These bacteria are prevalent in soil, water, and other organic debris-rich habitats.
- Examples: E. coli, Spirochaeta
Parasitic bacteria
- Parasitic bacteria are heterotrophic microorganisms that rely on living hosts for their nutritional requirements.
- They take nutrition from their host’s tissues or fluids by infecting other species such as plants, animals, or even other bacteria as hosts. Invading host cells, resisting host immune defenses, and obtaining nutrition for their own survival and reproduction are all strategies used by parasitic bacteria.
- The pathogens that cause illnesses including cholera, Lyme disease, and tuberculosis are examples of parasitic bacteria.
- Examples are Mycobacterium tuberculosis, Salmonella enterica
Anaerobic Heterotrophic Bacteria
Fermentative bacteria.
- A group of anaerobic heterotrophs known as fermentative bacteria use fermentation, a metabolic activity that enables them to produce energy in the absence of oxygen.
- These bacteria use organic substances as electron acceptors and donors to create molecules high in energy, such as adenosine triphosphate (ATP).
- Depending on the particular fermentative pathway used by the bacteria during fermentation, different end products, such as organic acids, alcohols, or gases, are frequently produced during the process.
- The gastrointestinal tracts of animals and the sediments of environments devoid of oxygen are only two examples of the many anaerobic habitats where fermentative bacteria can be found.
- Examples: Clostridium, Lactobacillus
Sulfate-Reducing Bacteria
- Anaerobic heterotrophic sulfate-reducing bacteria use sulfate (SO4-) as an electron acceptor during respiration.
- Because they transform sulfate into sulfide (S2), which can be used by other sulfur-oxidizing bacteria, these bacteria are essential to the sulfur cycle.
- Sulfate-reducing bacteria are frequently discovered in low oxygen environments, such as marine sediments, anaerobic sludge, or hydrothermal vents.
- Some sulfate-reducing microorganisms are also pathogenic, meaning they can harm both people and animals.
- Examples: Desulfovibrio, Desulfotomaculum
Mixotrophic bacteria
Autotrophic and heterotrophic type of metabolism are combined in a unique nutritional approach used by mixotrophic bacteria. These adaptable microbes can use both inorganic and organic carbon sources to meet their growth and energy requirements. Mixotrophic bacteria can adapt to changing environmental conditions and increase their chances of survival by switching between various nutritional modes.
Types of Mixotrophic Bacteria
Photoheterotrophic bacteria.
- Mixotrophic bacteria that can use light as an energy source but rely on organic carbon compounds as their main carbon source are known as photoheterotrophic bacteria.
- Similar to photosynthetic autotrophic bacteria, these bacteria use phototrophic pigments to gather light energy, but they also rely on organic molecules to obtain carbon and nutrients.
- Aquatic ecosystems are one setting where photoheterotrophic bacteria are common and where they contribute significantly to the carbon cycle.
- Examples: Rhodobacter, Rhodospirillum
Chemoheterotrophic Bacteria
- Chemoheterotrophic bacteria are mixotrophs that use organic carbon sources for growth and metabolism and obtain energy from the oxidation of organic molecules like sugars or organic acids.
- Due to their numerous metabolic pathways, these bacteria can convert complex organic molecules into simpler ones that can be used for energy production.
- Numerous ecosystems, including soil, water, and the microbiomes of plants and animals, are rich in chemoheterotrophic bacteria.
- Examples: Escherichia coli, Pseudomonas aeruginosa
Oligotrophic bacteria
Oligotrophic bacteria have developed particular characteristics to survive in conditions with less nutrients. They have mechanisms to scavenge and preserve vital nutrients, making them extremely effective at utilizing nutrients. In comparison to copiotrophic bacteria, oligotrophic bacteria frequently have smaller cell sizes, more streamlined genomes, and slower metabolic rates, which allow them to use scarce resources effectively.
Adaptations of Oligotrophic Bacteria to Low-Nutrient Environments
Oligotrophic bacteria use a variety of adaptations to survive in environments with few nutrients. These include the creation of specialized transport systems to gather limited nutrients, the synthesis of extracellular enzymes to effectively breakdown organic materials, and the construction of biofilms to improve nutrient uptake. To survive extended periods of food deprivation, oligotrophic bacteria can also create dormant forms, such as endospores.
Examples: Pelagibacter ubique, Prochlorococcus
Copiotrophic Bacteria,
Microorganisms known as copiotrophic bacteria grow in nutrient-rich habitats that are distinguished by a surplus of organic carbon and other nutrients. Compared to oligotrophic bacteria, they frequently have bigger cell sizes, better biomass yields, and more varied metabolic capabilities. Copiotrophic bacteria are frequently found in a variety of habitats, including rich soils, wastewater treatment facilities, and locations exposed to organic pollutants.
Adaptations of Copiotrophic Bacteria to High-Nutrient Environments
Copiotrophic bacteria have developed a variety of modifications to take advantage of situations rich in nutrients. They have developed regulatory mechanisms to quickly adapt to variations in nutrition availability, and they contain a variety of extracellular enzymes to effectively break down complicated chemical compounds. Copiotrophic bacteria perform crucial roles in the breakdown of organic matter and the cycling of nutrients. They have the ability to quickly colonize and dominate nutrient-rich settings.
Examples: Escherichia coli, Bacillus subtilis
Fastidious Bacteria
A group of microorganisms known as fastidious bacteria has extremely specific and demanding nutritional requirements. Fastidious bacteria have specialized nutritional requirements and frequently need sophisticated and enriched culture conditions in order to develop, unlike other bacteria that can grow on a variety of media. They are often fastidious in terms of pH, temperature, and oxygen conditions as well. Due to their unique niches or symbiotic relationships with other organisms, some bacteria may have evolved this dependency on particular nutrients.
Examples: Haemophilus influenzae (requires hemin and NAD), Neisseria gonorrhoeae (requires specific growth factors)
Symbiotic Microorganisms
Symbiotic bacteria interact in ways that can be advantageous or harmful for both organisms These other species can be other bacteria, plants, animals, or even other symbiotic bacteria. They frequently stay in certain host tissues or organs where they perform essential functions or take advantage of resources to ensure their own survival and reproduction. Symbiotic bacteria have developed ways to connect and communicate with their hosts, enabling them to form and sustain these relationships.
Mutualistic Symbiotic Bacteria
Plants with rhizobia and legumes.
Rhizobia are bacteria that produce nodules on the roots of legume plants and are mutualistic. Rhizobia in these nodules use nitrogen fixation to turn ambient nitrogen into ammonia, which the plants need as a source of nitrogen. Rhizobia receives nutrients and carbohydrates from the plants in exchange. Rhizobia may access a carbon supply that is rich in energy due to this mutualistic interaction, which also improves the growth and nitrogen nutrition of leguminous plants.
Examples: Rhizobium leguminosarum, Bradyrhizobium japonicum
Gut Microbiota and Humans
A diverse community of mutualistic bacteria that are present in the human gut are essential to the health of the host. These microorganisms help the body digest food, produce vitamins, metabolize bile acids, and control immunological function. Together, the gut microbiota and people have evolved over time, with the bacteria taking advantage of the gut’s nutrient-rich environment while the host received numerous advantages.
Examples: Bacteroides fragilis, Escherichia coli
Pathogenic Symbiotic Bacteria
Intracellular pathogens.
Some symbiotic bacteria can create intracellular infections in host cells, which can result in harmful effects. To enter host cells, avoid immune reactions, and take advantage of host resources, these bacteria have evolved sophisticated systems. Bacteria like Salmonella are examples of intracellular pathogens since they may enter host cells and multiply there, resulting in illnesses like typhoid fever.
Examples: Mycobacterium tuberculosis, Salmonella enterica
Extracellular Pathogens
extracellular symbiotic bacteria can cause disease By colonization and multiplying in host tissues or bodily fluids. They develop virulence factors that give them the ability to get across the host’s defenses and damage the tissues of the host. Bacteria like Streptococcus pneumoniae, which may cause pneumonia and other illnesses, are an example of an extracellular pathogen.
Examples: Streptococcus pneumoniae, Staphylococcus aureus
Commensal Bacteria
Bacteria known as commensal bacteria survive with other organisms without causing harm or giving obvious advantages. These microorganisms live inside their hosts in a variety of ecological niches, frequently colonizing bodily surfaces including the skin, mouth cavity, or gastrointestinal system.
Examples: Staphylococcus epidermidis, Bifidobacterium spp.
Amensalistic Bacteria
Amensalistic bacteria interact in a way that causes one organism to suffer while leaving the other unharmed. In these interactions, one organism releases substances or elements that prevent or reduce the development or survival of another organism.
Antibiotics or toxins that amensalistic bacteria secrete prevent the growth or survival of other species without providing any direct benefit to themselves. These interactions are frequently competitive in nature, giving the bacteria an advantage over their competitors by lowering the fitness or viability of nearby organisms.
Examples of Amensalistic Interactions between Bacteria and Other Organisms
There are many environments where bacteria and other species interact amensally. For example, several soil bacteria create antibiotics that prevent the expansion of neighboring microbes, reducing competition for resources. Similar to bacteriocins, which are antimicrobial peptides that inhibit the growth of potential pathogens and provide a protective impact on the host, some bacteria in the human microbiota also generate them.
Examples: Streptomyces spp., Penicillium spp.
In conclusion, bacterial nutrition is a complex and diverse field that encompasses a wide array of metabolic strategies and ecological interactions. Autotrophic bacteria harness energy from inorganic sources, while heterotrophic bacteria rely on organic compounds. Mixotrophic bacteria combine both autotrophic and heterotrophic modes of nutrition
- Tortora, G. J., Funke, B. R., & Case, C. L. (2021). Microbiology: An introduction. Pearson Education Limited. Willey, J. M., Sandman, K. M., Wood, D. H., & Prescott, L. M. (2019). Prescott’s microbiology (11th ed.). McGraw Hill.
- Greemwood, R. S. (2002). Medical Microbiology. London: Churchill Livingstone.
- J.Pelezar. (1993). Microbiology. Tata McGraw hill.
- https://www.ncbi.nlm.nih.gov/books/NBK8406/

Mubashir Iqbal
Mubashir Iqbal is a highly dedicated and motivated Microbiologist with an MPhil in Microbiology from the University of Veterinary and Animal Sciences. Currently, he is researching the efficacy of commercially available SARS Cov-2 vaccines to neutralize the omicron variant in Pakistan. He holds a Bachelor's degree in Microbiology and has experience in chemical and microbiological analysis of water samples, managing SOPs and documents according to standard ISO 17025. Additionally, he has worked as an internee in BSL 3, Institute of Microbiology, UVAS, where he gained experience in RNA extraction, sample processing, and microscopy.
Newsletter Updates
Enter your email address below and subscribe to our newsletter
I accept the Privacy Policy
Leave a Reply Cancel Reply
Your email address will not be published. Required fields are marked *
Add Comment *
Save my name, email, and website in this browser for the next time I comment.
Post Comment
Related Posts

Isolation and Identification of Salmonella typhi in easy steps
- April 21, 2024

Isolation and Identification of Staphylococcus aureus
- February 22, 2024

Acetamide Utilization Test – An overview
- February 19, 2024
Trending now

11 Microbial Nutrition
Nutritional types of microorganisms.
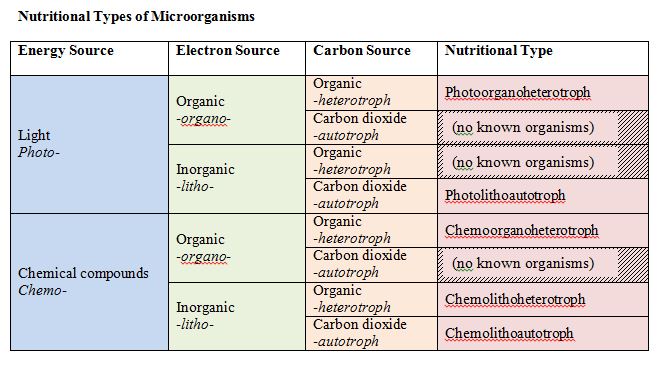
All microbes have a need for three things: carbon, energy, and electrons. There are specific terms associated with the source of each of these items, to help define organisms.
Let us focus on carbon first. All organisms are carbon-based with macromolecules – proteins, carbohydrates, lipids, nucleic acid – having a fundamental core of carbon. On one hand, organisms can use reduced, preformed organic substances as a carbon source. These are the heterotrophs or “other eaters.” Alternatively, they can rely on carbon dioxide (CO2) as a carbon source, reducing or “fixing” it this inorganic form of carbon into an organic molecule. These are the autotrophs or “self feeders.”
For energy, there are two possibilities as well: light energy or chemical energy. Light energy comes from the sun, while chemical energy can come from either organic or inorganic chemicals. Those organisms that use light energy are called phototrophs (“light eaters”), while those that use chemical energy are called chemotrophs (“chemical eaters”). Chemical energy can come from inorganic sources or organic sources. An organism that uses inorganic sources is known as a lithotroph (“rock eater”), while an organism that uses organic sources is called an organotroph (“organic eater”).
These terms can all be combined, to derive a single term that gives you an idea of what an organism is using to meet its basic needs for energy, electrons, and carbon.
Macronutrients
In addition to carbon, hydrogen and oxygen, cells need a few other elements in sufficient quantity. In particular, cells need nitrogen for the formation of proteins, nucleic acids, and a few other cell components. Cells also need phosphorous, which is a crucial component of nucleic acids (think sugar- phosphate backbone!), phospholipids, and adenosine tri phosphate or ATP. Sulfur is necessary for a few amino acids, as well as several vitamins, while potassium is needed for enzymes, and magnesium is used to stabilize ribosomes and membrane. Collectively these elements (including C, H, and O) are referred to as the macronutrients .
Growth Factors
Some microbes can synthesize certain organic molecules that they need from scratch, as long as they are provided with carbon source and inorganic salts. Other microbes require that certain organic compounds exist within their environment. These organic molecules essential for growth are called growth factors and fall in three categories: 1) amino acids (building blocks of protein), 2) purines and pyrimidines (building blocks of nucleic acid), and 3) vitamins (enzyme cofactors).
Uptake of Nutrients
In order to support its’ activities, a cell must bring in nutrients from the external environment across the cell membrane. In bacteria and archaea, several different transport mechanisms exist.
Passive Diffusion
Passive or simple diffusion allows for the passage across the cell membrane of simple molecules and gases, such as CO2, O2, and H2O. In this case, a concentration gradient must exist, where there is higher concentration of the substance outside of the cell than there is inside the cell. As more of the substance is transported into the cell the concentration gradient decreases, slowing the rate of diffusion.
Facilitated Diffusion
Facilitated diffusion also involves the use of a concentration gradient, where the concentration of the substance is higher outside the cell, but differs with the use of carrier proteins (sometimes called permeases ). These proteins are embedded within the cell membrane and provide a channel or pore across the membrane barrier, allowing for the passage of larger molecules. If the concentration gradient dissipates, the passage of molecules into the cell stops. Each carrier protein typically exhibits specificity, only transporting in a particular type of molecule or closely related molecules.
Active Transport
Many types of nutrient uptake require that a cell be able to transport substances against a concentration gradient (i.e., with a higher concentration inside the cell than outside). In order to do this, a cell must utilize metabolic energy for the transport of the substance through carrier proteins embedded in the membrane. This is known as active transport . All types of active transport utilize carrier proteins.

There are three main examples of active transport: primary active transport, secondary active transport, and group translocation.
Primary active transport
Primary active transport involves the use of chemical energy, such as ATP, to drive the transport. One example is the ABC system , which utilizes ATP-Binding Cassette transporters . Each ABC transporter is composed of three different components: 1) membrane-spanning proteins that form a pore across the cell membrane (i.e. carrier protein), 2) an ATP binding region that hydrolyzes ATP, providing the energy for the passage across the membrane, and 3) a substrate-binding protein, a peripheral protein that binds to the appropriate substance to be transporter and ferries it to the membrane-spanning proteins. In gram negative bacteria the substrate-binding protein is located in the cell’s periplasm, while in gram positive bacteria the substrate-binding protein is attached to the outside of the cell membrane.

Secondary active transport
Secondary active transport utilizes energy from a proton motive force (PMF) . A PMF is an ion gradient that develops when the cell transports electrons during energy-conserving processes. Positively charged protons accumulate along the outside of the negatively charged cell, creating a proton gradient between the outside of the cell and the inside.
There are three different types of transport events for simple transport: uniport , symport , and antiport and each mechanism utilizes a different protein porter . Uniporters transport a single substance across the membrane, either in or out. Symporters transport two substances across the membrane at the same time, typically a proton paired with another molecule. Antiporters transport two substances across the membrane as well, but in opposite directions. As one substance enters the cell, the other substance is transported out.

Group Translocation
Group translocation is a distinct type of active transport, using energy from an energy-rich organic compound that is not ATP. Group translocation also differs from both simple transport and ABC transporters in that the substance being transported is chemically modified in the process.
One of the best studied examples of group translocation is the phosphoenolpyruvate: sugar phosphotransferase system (PTS) , which uses energy from the high-energy molecule phosphoenolpyruvate (PEP) to transport sugars into the cell. A phosphate is transferred from the PEP to the incoming sugar during the process of transportation.

Iron Uptake

Iron is required by microbes for the function of their cytochromes and enzymes, resulting in it being a growth-limiting micronutrient. However, little free iron is available in environments, due to its insolubility. Many bacteria have evolved siderophores , organic molecules that chelate or bind ferric iron with high affinity. Siderophores are released by the organism to the surrounding environment, whereby they bind any available ferric iron. The iron-siderophore complex is then bound by a specific receptor on the outside of the cell, allowing the iron to be transported into the cell.
heterotroph, autotroph, phototroph, chemotroph, lithotroph, organotroph, photolithoautotroph, photoorganoheterotroph, chemoorganoheterotroph, chemolithoautotroph, chemolithoheterotroph, macronutrients, growth factors, passive/simple diffusion, facilitated diffusion, carrier protein/permease, active transport, primary active transport, ABC system, ATP-binding cassette transporter, ABC transporter, secondary active transport, proton motive force (PMF), uniport, symport, antiport, porter, uniporter, symporter, antiporter, group translocation, phosphoenolpyruvate: sugar phosphotransferase system (PTS), phosphoenolpyruvate (PEP), siderophore.
Study Questions
- What are the different terms associated with microbial nutritional types? How can these terms be combined to define the nutritional types of microbes in terms of their sources of carbon, electrons, and energy?
- What are macroelements and why are they important to a cell? What are growth factors and what is their significance to a cell?
- What is the importance of nutrient uptake for a cell? What are the common features of nutrient uptake by bacteria?
- What is transported into a bacteria cell by passive diffusion and how does this affect a bacterial cell?
- Explain diffusion (passive and facilitated) and active transport.
- What are the 3 types of active transport? Be able to diagram each processes. What is required for each of these processes? How are they similar, how are they different?
- Why is iron uptake important for a cell? What is used to accomplish this?
Exploratory Questions (OPTIONAL)
- What is the largest bacterium or archaean ever discovered? What is the smallest eukaryote ever discovered?
General Microbiology Copyright © 2019 by Linda Bruslind is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License , except where otherwise noted.

- school Campus Bookshelves
- menu_book Bookshelves
- perm_media Learning Objects
- login Login
- how_to_reg Request Instructor Account
- hub Instructor Commons
Margin Size
- Download Page (PDF)
- Download Full Book (PDF)
- Periodic Table
- Physics Constants
- Scientific Calculator
- Reference & Cite
- Tools expand_more
- Readability
selected template will load here
This action is not available.
5.3: Bacteria Nutrition
- Last updated
- Save as PDF
- Page ID 2939
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)

Can bacteria make their own food from sunlight?
Plants aren't the only organisms that use the energy of the sun to make food. Some bacteria can also perform photosynthesis. In fact, the first photosynthetic organisms on Earth were bacteria. Photosynthesis is just one of many ways that bacteria can obtain energy.
Bacteria Nutrition
Like all organisms, bacteria need energy, and they can acquire this energy through a number of different ways.
Photosynthesis
Photosynthetic bacteria use the energy of the sun to make their own food. In the presence of sunlight, carbon dioxide and water are turned into glucose and oxygen. The glucose is then turned into usable energy. Glucose is like the "food" for the bacteria. An example of photosynthetic bacteria is cyanobacteria, as seen in the opening image. These bacteria are sometimes called blue-green algae, although they are not algae, due to their numerous chlorophyll molecules.
Decomposers
Bacteria known as decomposers break down wastes and dead organisms into smaller molecules. These bacteria use the organic substrates they break down to get their energy, carbon, and nutrients they need for survival.
Chemotrophs
Bacteria can also be chemotrophs. Chemosynthetic bacteria, or chemotrophs , obtain energy by breaking down chemical compounds in their environment. An example of one of these chemicals broken down by bacteria is nitrogen-containing ammonia. These bacteria are important because they help cycle nitrogen through the environment for other living things to use. Nitrogen cannot be made by living organisms, so it must be continually recycled. Organisms need nitrogen to make organic compounds, such as DNA.
Some bacteria depend on other organisms for survival. For example, some bacteria live in the roots of legumes, such as pea plants (Figure below). The bacteria turn nitrogen-containing molecules into nitrogen that the plant can use. Meanwhile, the root provides nutrients to the bacteria. In this relationship, both the bacteria and the plant benefit, also known as mutualism .
Other mutualistic bacteria include gut microbes. These are bacteria that live in the intestines of animals . They are usually beneficial bacteria, needed by the host organism. These microbes obviously don't kill their host, as that would kill the bacteria as well.

Other bacteria are parasitic and can cause illness. In parasitism , the bacteria benefit, and the other organism is harmed. Harmful bacteria will be discussed in another concept.
- Bacteria can obtain energy and nutrients by performing photosynthesis, decomposing dead organisms and wastes, or breaking down chemical compounds.
- Bacteria can obtain energy and nutrients by establishing close relationships with other organisms, including mutualistic and parasitic relationships.
Explore More
Use the resources below to answer the questions that follow.
Explore More I
- Bacteria http://www.youtube.com/watch?v=h-z9-9OOWC4 (11:04)
- What are the three nutritional types of bacteria?
- What is the base energy source for these three types?
- Give an example of a photoautotroph.
Explore More II
- Bacteria: Life History and Ecology http://www.ucmp.berkeley.edu/bacteria/bacterialh.html
- What is the difference between a heterotroph and an autotroph?
- What is the difference between aerobic, anaerobic, and facultative anaerobic bacteria?
- What are two ways bacteria play important roles in the ecosystem?
- Describe two ways that bacteria obtain nutrients and energy?
- What is an example of a mutualism with a bacteria?
- What is an example of a photosynthetic bacteria.
- Describe the importance of chemosynthetic bacteria.
Nutritional Types of Bacteria
All organisms require an energy source to drive energy-consuming life processes. Energy can be tapped from light or chemicals (organic chemicals and inorganic chemicals). The nutritional types of bacteria vary greatly. Organisms that can utilize radiant energy (sunlight) are called phototrophs. Chemotrophs are organisms that can harvest energy from chemicals.
Table of Contents
Chemotrophs
Chemoorganotrophs.
Organisms that conserve energy from organic chemicals are called chemoorganotrophs . Thousands of different organic chemicals are nutritional types and can be used by one or another microorganism. Indeed, all-natural and even most synthetic organic compounds can be metabolized. Energy is conserved from the oxidation of the compound and is stored in the cell in the energy-rich bonds of the compound adenosine triphosphate (ATP).
Aerobes obtain energy from an organic compound in the presence of oxygen, anaerobes obtain energy in the absence of oxygen and facultative anaerobes can break down organic compounds in both aerobic and anaerobic conditions.

Chemolithotrophs
The oxidation of inorganic compounds to yield energy is known as chemolithotrophy . Many prokaryotes can tap the energy available from the oxidation of inorganic compounds. This phenomenon was discovered by the Russian microbiologist Winogradsky. Organisms that carry out chemolithotrophic reactions are called chemolithotrophs . Like phototrophic organisms, chemolithotrophic bacteria are also autotrophs.
Chemolithotrophy occurs only in prokaryotes and is widely distributed among species of Bacteria and Archaea . Several inorganic compounds can be oxidized; for example, H2, H2S (hydrogen sulfide), NH3 (ammonia), and Fe21 (ferrous iron). Typically, a related group of chemolithotrophs specializes in the oxidation of a related group of inorganic compounds, and thus we have the “sulfur” bacteria, the “iron” bacteria, and so on.
The capacity to conserve energy from the oxidation of inorganic chemicals is a good metabolic strategy because competition from chemoorganotrophs, organisms that require organic energy sources, is not an issue. In addition, many of the inorganic compounds oxidized by chemolithotrophs, for example, H2 and H2S, are actually the waste products of chemoorganotrophs. Thus, chemolithotrophs have evolved strategies for exploiting resources that chemoorganotrophs are unable to use, so it is common for species of these two physiological groups to live in close association with one another.
Phototrophs
Sunlight is available in many microbial habitats on Earth, phototrophic microorganisms living in those areas harvest energy from sunlight. They contain pigments that allow them to convert light energy into chemical energy, and thus their cells appear colored. Unlike chemotrophic organisms, phototrophs do not require chemicals as a source of energy.
Purple bacteria appeared on Earth long before oxygenic phototrophs evolved. Green sulfur bacteria were some of the first phototrophs to evolve on Earth.
Two major forms of phototrophy are known in prokaryotes.
- Oxygenic photosynthesis : oxygen (O2) is produced. Among microorganisms, oxygenic photosynthesis is characteristic of cyanobacteria and algae (oxygenic phototrops).
- Anoxygenic photosynthesis : does not yield O2. Purple sulfur bacteria, green bacteria, and heliobacteria are anoxygenic phototrophs.

Photolithotrophs
Among phototrophic bacteria are species that use inorganic compounds as their source of electrons and are called photolithotrophs . For example, Chromatium okenii.
Photoorganotrophs
Some phototrophic microorganisms use organic compounds such as fatty acids and alcohols as electron donors and are therefore photoorganotrophs . For example, Rhodospirillum rubrum.
Heterotrophs and Autotrophs
All organisms require carbon in some form either in small or large amounts to synthesize cell components. Organisms that can use carbon dioxide (CO2) as their major or even sole source of carbon are termed autotrophs. Other organisms require organic compounds as their carbon source and are known as heterotrophs.
Chemoorganotrophs are by definition heterotrophs. By contrast, most chemolithotrophs and phototrophs are autotrophs. For example chemolithotrophic bacteria of the genus Nitrosomonas are able to oxidize ammonia into nitrite, thereby obtaining sufficient energy to assimilate the carbon of CO2 into cell component (CO2 fixation).
These are sometimes called primary producers because they synthesize new organic matter from CO2 for both their own benefit and that of chemoorganotrophs. Autotrophs can transform inorganic compounds into carbohydrates, proteins, nucleic acids, lipids, vitamins, and other complex organic substances required for the cells.
Heterotrophs either feed directly on the cells of primary producers or live off products they excrete. Virtually all organic matter on Earth has been synthesized by primary producers, in particular, the phototrophs. Autotrophs are responsible for the cycling of elements in nature through biological processes.
Heterotrophs
Heterotrophs rely on autotrophs for their foods and are also called consumers of the food chains. All the organisms that cause diseases of humans, animals, and plants are heterotrophs. They constitute the greater part of the microbial population in our immediate environment. Heterotrophs vary considerably in their nutritional requirements, particularly with respect to their organic carbon source, nitrogen sources, and vitamins. For example, E. coli has simple nutritional requirements than lactobacilli.
Obligate Parasites
The nutritional and physical requirements of this group of bacteria are not known yet so we can not cultivate them in an artificial medium. Such bacteria are propagated only through animal inoculation. For example, Mycobacterium leprae can be cultured by infection suckling mice or nine-banded armadillos. Other obligate intracellular bacteria are rickettsia, chlamydias, and spirochetes.
References and further readings
- Madigan Michael T, Bender, Kelly S, Buckley, Daniel H, Sattley, W. Matthew, & Stahl, David A. (2018). Brock Biology of Microorganisms (15th Edition). Pearson.
- Pelczar, M. J., Chan, E. C. S., & Krieg, N. R. (2001). Microbiology: Concepts and applications. New York: McGraw-Hill.
Acharya Tankeshwar
Hello, thank you for visiting my blog. I am Tankeshwar Acharya. Blogging is my passion. As an asst. professor, I am teaching microbiology and immunology to medical and nursing students at PAHS, Nepal. I have been working as a microbiologist at Patan hospital for more than 10 years.
We love to get your feedback. Share your queries or comments Cancel reply
This site uses Akismet to reduce spam. Learn how your comment data is processed .
Recent Posts
Urea Cycle: Steps, End Products, and Functions
Amino nitrogen, a key component in the synthesis of amino acids or new nitrogenous products, can be toxic to the human body if not utilized to create new compounds. To prevent this, ureotelic...
Gluconeogenesis: Enzymes Involved, Steps, and Functions
During fasting, vigorous exercise, and hypoglycemic conditions, the body requires high glucose. Gluconeogenesis converts non-carbohydrate molecules like glycerol, pyruvate, lactate, glucogenic amino...

- Biology Article
One of the very first organisms to evolve on earth was probably a unicellular organism, similar to modern bacteria. Ever since then, life has evolved into a multitude of life forms over many millennia. However, we can still trace our ancestry back to this single-celled organism.
Table of Contents
Bacteria Diagram
- Ultrastructure of a Bacterial Cell
- Classification
- Reproduction
Useful Bacteria
Harmful bacteria.
Today, bacteria are considered as one of the oldest forms of life on earth. Even though most bacteria make us ill, they have a long-term, mutual relationship with humans and are very much important for our survival. But before we elaborate on its uses, let us know the structure of bacteria, its classification, and the bacteria diagram in detail.
Bacteria Definition
“Bacteria are unicellular organisms belonging to the prokaryotic group where the organisms lack a few organelles and a true nucleus”.
Also Read: Gram Negative Bacteria
The bacteria diagram given below represents the structure of a typical bacterial cell with its different parts. The cell wall, plasmid, cytoplasm and flagella are clearly marked in the diagram.

Bacteria Diagram representing the Structure of Bacteria
Ultrastructure of a Bacteria Cell
The structure of bacteria is known for its simple body design. Bacteria are single-celled microorganisms with the absence of the nucleus and other c ell organelles ; hence, they are classified as prokaryotic organisms.
They are also very versatile organisms, surviving in extremely inhospitable conditions. Such organisms are called extremophiles. Extremophiles are further categorized into various types based on the types of environments they inhabit:
- Thermophiles
- Acidophiles
- Alkaliphiles
Another fascinating feature of bacteria is their protective cell wall , which is made up of a special protein called peptidoglycan. The components of bacterial cell wall forms an important basis upon which the bacteria can be divided. This particular protein isn’t found anywhere else in nature except in the cell walls of bacteria.
But few of them are devoid of this cell wall, and others have a third protection layer called capsule. On the outer layer, one or more flagella or pili is attached, and it functions as a locomotory organ. Pili can also help certain bacteria to attach themselves to the host’s cells. They do not contain any cell organelle as in animal or plant cell except for ribosomes.
Ribosomes are the sites of protein synthesis. In addition to this DNA, they have an extra circular DNA called plasmid. These plasmids make some strains of bacteria resistant to antibiotics.
Also Read: Gram Positive Bacteria
Classification of Bacteria
Bacteria can be classified into various categories based on their features and characteristics. The classification of bacteria is mainly based on the following:
- Composition of the cell wall
- Mode of respiration
- Mode of nutrition
Also check: Bergey’s Classification of Bacteria
Classification of bacteria based on Shape
Classification of bacteria based on the composition of the cell wall, classification of bacteria based on the mode of nutrition, classification of bacteria based on the mode of respiration.
Also Read: Difference between Bacteria and Virus
Reproduction in Bacteria

Bacteria follow an asexual mode of reproduction , called binary fission. A single bacterium divides into two daughter cells. These are identical to the parent cell as well as to each other. Replication of DNA within the parent bacterium marks the beginning of the fission. Eventually, cell elongates to form two daughter cells.
The rate and timing of reproduction depend upon the conditions like temperature and availability of nutrients. When there is a favourable condition, E.coli or Escherichia coli produces about 2 million bacteria every 7 hours.
Bacterial reproduction is strictly asexual, but it can undergo sexual reproduction in very rare cases.
Genetic recombination in bacteria has the potential to occur through conjugation, transformation, or transduction. In such cases, the bacteria may become resistant to antibiotics since there is variation in the genetic material (as opposed to asexual reproduction where the same genetic material is present in generations)
Also Read: Binary fission
Not all bacteria are harmful to humans. There are some bacteria which are beneficial in different ways. Listed below are few benefits of bacteria:
- Convert milk into curd – Lactobacillus or lactic acid bacteria
- Ferment food products – Streptococcus and Bacillus
- Help in digestion and improving the body’s immunity system – Actinobacteria, Bacteroidetes, Firmicutes, Proteobacteria
- Production of antibiotics, which is used in the treatment and prevention of bacterial infections – Soil bacteria
Also Refer: Antibiotics
There are bacteria that can cause a multitude of illnesses. They are responsible for many of the i nfectious diseases like pneumonia, tuberculosis, diphtheria, syphilis, tooth decay. Their effects can be rectified by taking antibiotics and prescribed medication.
However, precaution is much more effective. Most of these disease-causing bacteria can be eliminated by sterilizing or disinfecting exposed surfaces, instruments, tools and other utilities. These methods include- application of heat, disinfectants, UV radiations, pasteurization, boiling, etc.
To know more about bacteria, its definition, the structure of bacteria, bacteria diagram, classification of bacteria, and reproduction in bacteria keep visiting BYJU’S website or download BYJU’S app for further reference.

- Cryptobiosis
- Microorganisms: Friend And Foe
Frequently Asked Questions
1. what are the different types of bacteria.
Bacteria can be divided into several types based on several characteristics such as shape, cell wall composition, mode of respiration, and mode of nutrition.
2. What is bacteria? How do you define bacteria?
Bacteria are prokaryotic unicellular organisms. They have a relatively simple cell structure compared to eukaryotic cells. They also do not possess any membrane-bound organelles such as a nucleus. However, do they possess genetic material (DNA or RNA) in the intracellular space called the nucleoid
3. How do bacteria reproduce?
Bacteria reproduce through a process called binary fission. In this process, a single bacterium divides into two daughter cells. These daughter cells are identical to the parent cell as well as to each other.
4. State 4 examples of bacteria.
- Streptococcus
- Actinobacteria
- Proteobacteria
5. The study of bacteria is called?
The study of bacteria is called bacteriology.
6. What are the examples of acidophilic bacteria?
Acetobacter aceti and Alicyclobacillus acidiphilus are two examples of acidophilic bacteria.
Also check:
- Bacterial Cell Division
- Composition of Bacterial Cell Wall
- Classification of Culture Media
- evolution of microbiology

Put your understanding of this concept to test by answering a few MCQs. Click ‘Start Quiz’ to begin!
Select the correct answer and click on the “Finish” button Check your score and answers at the end of the quiz
Visit BYJU’S for all Biology related queries and study materials
Your result is as below
Request OTP on Voice Call
Leave a Comment Cancel reply
Your Mobile number and Email id will not be published. Required fields are marked *
Post My Comment
please state about eubacteria and archeabacteria
Read more about eubacteria and archaebacteria here .
Thank you so much byjus . This helped me a lot
Good Information Thanks to BYJU’S
THANK YOU .
Please tell me about kinds of Bacteria
Bacteria can be classified based on shape, mode of nutrition, respiration, the composition of the cell wall, etc. Based on these criteria, bacteria can be classified as bacillus, coccus or vibrio, etc., autotrophic or heterotrophic, aerobic or anaerobic, Gram + or Gram -. Read the above topic for detailed information.
Good info it’s REALLY helpful in my exam
Difference between Bacteria and Virus
Classify bacteria on the basic of oxygen requirements?
Yes it is really good 👌👌
- Share Share
Register with BYJU'S & Download Free PDFs
Register with byju's & watch live videos.


- school Campus Bookshelves
- menu_book Bookshelves
- perm_media Learning Objects
- login Login
- how_to_reg Request Instructor Account
- hub Instructor Commons
Margin Size
- Download Page (PDF)
- Download Full Book (PDF)
- Periodic Table
- Physics Constants
- Scientific Calculator
- Reference & Cite
- Tools expand_more
- Readability
selected template will load here
This action is not available.
4.2: Microbial Nutrition
- Last updated
- Save as PDF
- Page ID 86510

- Linda Bruslind
- Oregon State University via Open Oregon State
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
All microbes have a need for three things: carbon, energy, and electrons. There are specific terms associated with the source of each of these items, to help define organisms.
Let us focus on carbon first. All organisms are carbon-based with macromolecules – proteins, carbohydrates, lipids, nucleic acid – having a fundamental core of carbon. On one hand, organisms can use reduced, preformed organic substances as a carbon source. These are the heterotrophs or “other eaters.” Alternatively, they can rely on carbon dioxide (CO2) as a carbon source, reducing or “fixing” it this inorganic form of carbon into an organic molecule. These are the autotrophs or “self feeders.”
For energy, there are two possibilities as well: light energy or chemical energy. Light energy comes from the sun, while chemical energy can come from either organic or inorganic chemicals. Those organisms that use light energy are called phototrophs (“light eaters”), while those that use chemical energy are called chemotrophs (“chemical eaters”). Chemical energy can come from inorganic sources or organic sources. An organism that uses inorganic sources is known as a lithotroph (“rock eater”), while an organism that uses organic sources is called an organotroph (“organic eater”).
These terms can all be combined, to derive a single term that gives you an idea of what an organism is using to meet its basic needs for energy, electrons, and carbon.

Macronutrients
In addition to carbon, hydrogen and oxygen, cells need a few other elements in sufficient quantity. In particular, cells need nitrogen for the formation of proteins, nucleic acids, and a few other cell components. Cells also need phosphorous, which is a crucial component of nucleic acids (think sugar- phosphate backbone!), phospholipids, and adenosine tri phosphate or ATP. Sulfur is necessary for a few amino acids, as well as several vitamins, while potassium is needed for enzymes, and magnesium is used to stabilize ribosomes and membrane. Collectively these elements (including C, H, and O) are referred to as the macronutrients .
Growth Factors
Some microbes can synthesize certain organic molecules that they need from scratch, as long as they are provided with carbon source and inorganic salts. Other microbes require that certain organic compounds exist within their environment. These organic molecules essential for growth are called growth factors and fall in three categories: 1) amino acids (building blocks of protein), 2) purines and pyrimidines (building blocks of nucleic acid), and 3) vitamins (enzyme cofactors).
Uptake of Nutrients
In order to support its’ activities, a cell must bring in nutrients from the external environment across the cell membrane. In bacteria and archaea, several different transport mechanisms exist.
Passive Diffusion
Passive or simple diffusion allows for the passage across the cell membrane of simple molecules and gases, such as CO2, O2, and H2O. In this case, a concentration gradient must exist, where there is higher concentration of the substance outside of the cell than there is inside the cell. As more of the substance is transported into the cell the concentration gradient decreases, slowing the rate of diffusion.
Facilitated Diffusion
Facilitated diffusion also involves the use of a concentration gradient, where the concentration of the substance is higher outside the cell, but differs with the use of carrier proteins (sometimes called permeases ). These proteins are embedded within the cell membrane and provide a channel or pore across the membrane barrier, allowing for the passage of larger molecules. If the concentration gradient dissipates, the passage of molecules into the cell stops. Each carrier protein typically exhibits specificity, only transporting in a particular type of molecule or closely related molecules.
Active Transport
Many types of nutrient uptake require that a cell be able to transport substances against a concentration gradient (i.e. with a higher concentration inside the cell than outside). In order to do this, a cell must utilize metabolic energy for the transport of the substance through carrier proteins embedded in the membrane. This is known as active transport . All types of active transport utilize carrier proteins.

Active Transport Versus Facilitated Diffusion.
Primary active transport
Primary active transport involves the use of chemical energy, such as ATP, to drive the transport. One example is the ABC system , which utilizes ATP-Binding Cassette transporters . Each ABC transporter is composed of three different components: 1) membrane-spanning proteins that form a pore across the cell membrane (i.e. carrier protein), 2) an ATP binding region that hydrolyzes ATP, providing the energy for the passage across the membrane, and 3) a substrate-binding protein, a peripheral protein that binds to the appropriate substance to be transporter and ferries it to the membrane-spanning proteins. In gram negative bacteria the substrate-binding protein is located in the cell’s periplasm, while in gram positive bacteria the substrate-binding protein is attached to the outside of the cell membrane.

ABC Transporter Structure.
Secondary active transport
Secondary active transport utilizes energy from a proton motive force (PMF) . A PMF is an ion gradient that develops when the cell transports electrons during energy-conserving processes. Positively charged protons accumulate along the outside of the negatively charged cell, creating a proton gradient between the outside of the cell and the inside.
There are three different types of transport events for simple transport: uniport , symport , and antiport and each mechanism utilizes a different protein porter . Uniporters transport a single substance across the membrane, either in or out. Symporters transport two substances across the membrane at the same time, typically a proton paired with another molecule. Antiporters transport two substances across the membrane as well, but in opposite directions. As one substance enters the cell, the other substance is transported out.

Uniport Synport Antiport. By Lupask (Own work) [Public domain], via Wikimedia Commons
Group Translocation
Group translocation is a distinct type of active transport, using energy from an energy-rich organic compound that is not ATP. Group translocation also differs from both simple transport and ABC transporters in that the substance being transported is chemically modified in the process.
One of the best studied examples of group translocation is the phosphoenolpyruvate: sugar phosphotransferase system (PTS) , which uses energy from the high-energy molecule phosphoenolpyruvate (PEP) to transport sugars into the cell. A phosphate is transferred from the PEP to the incoming sugar during the process of transportation.

Group Translocation via PTS.
Iron Uptake
Iron is required by microbes for the function of their cytochromes and enzymes, resulting in it being a growth-limiting micronutrient. However, little free iron is available in environments, due to its insolubility. Many bacteria have evolved siderophores , organic molecules that chelate or bind ferric iron with high affinity. Siderophores are released by the organism to the surrounding environment, whereby they bind any available ferric iron. The iron-siderophore complex is then bound by a specific receptor on the outside of the cell, allowing the iron to be transported into the cell.

Siderophores and Receptor Sites.
heterotroph, autotroph, phototroph, chemotroph, lithotroph, organotroph, photolithoautotroph, photoorganoheterotroph, chemoorganoheterotroph, chemolithoautotroph, chemolithoheterotroph, macronutrients, growth factors, passive/simple diffusion, facilitated diffusion, carrier protein/permease, active transport, primary active transport, ABC system, ATP-binding cassette transporter, ABC transporter, secondary active transport, proton motive force (PMF), uniport, symport, antiport, porter, uniporter, symporter, antiporter, group translocation, phosphoenolpyruvate: sugar phosphotransferase system (PTS), phosphoenolpyruvate (PEP), siderophore.
Study Questions
- What are the different terms associated with microbial nutritional types? How can these terms be combined to define the nutritional types of microbes in terms of their sources of carbon, electrons, and energy?
- What are macroelements and why are they important to a cell? What are growth factors and what is their significance to a cell?
- What is the importance of nutrient uptake for a cell? What are the common features of nutrient uptake by bacteria?
- What is transported into a bacteria cell by passive diffusion and how does this affect a bacterial cell?
- Explain diffusion (passive and facilitated) and active transport.
- What are the 3 types of active transport? Be able to diagram each processes. What is required for each of these processes? How are they similar, how are they different?
- Why is iron uptake important for a cell? What is used to accomplish this?
Exploratory Questions (OPTIONAL)
- What is the largest bacterium or archaean ever discovered? What is the smallest eukaryote ever discovered?

An official website of the United States government
The .gov means it’s official. Federal government websites often end in .gov or .mil. Before sharing sensitive information, make sure you’re on a federal government site.
The site is secure. The https:// ensures that you are connecting to the official website and that any information you provide is encrypted and transmitted securely.
- Publications
- Account settings
Preview improvements coming to the PMC website in October 2024. Learn More or Try it out now .
- Advanced Search
- Journal List
- Elsevier - PMC COVID-19 Collection
Classification, identification and typing of micro-organisms
- • Taxonomy is the classification, nomenclature and identification of microbes (algae, protozoa, slime moulds, fungi, bacteria, archaea and viruses). The naming of organisms by genus and species is governed by an international code.
- • Bacteria can be separated into two major divisions by their reaction to Gram's stain, and exhibit a range of shapes and sizes from spherical (cocci) through rod shaped (bacilli) to filaments and spiral shapes.
- • In clinical practice, bacteria are classified by macroscopic and microscopic morphology, their requirement for oxygen, and activity in phenotypic and biochemical tests.
- • Various diagnostic test systems are used to detect specific bacteria in clinical systems, including specific gene probes, reaction with antibodies in ELISA formats, immunofluorescence and, increasingly, PCR-based technology.
- • Different bacterial species often exhibit different population structures, highly diverse (panmictic) or relatively uniform (clonal) depending mainly on the frequency of gene recombination (from external sources).
- • Typing of bacterial isolates is necessary for epidemiological investigations in outbreaks and for surveillance, and a variety of phenotypic and genetic methods has evolved for the identification of strains.
Micro-organisms may be classified in the following large biological groups:
- 3 Slime moulds
The algae (excluding the blue–green algae), the protozoa, slime moulds and fungi include the larger eukaryotic (see Ch. 2) micro-organisms; their cells have the same general type of structure and organization as that found in plants and animals. The bacteria, including organisms of the mycoplasma, rickettsia and chlamydia groups, together with the related blue–green algae, comprise the smaller micro-organisms, with the form of cellular organization described as prokaryotic . The archaea are a distinct phylogenetic group of prokaryotes that bear only a remote ancestral relationship to other organisms (see Ch. 2). As the algae, slime moulds and archaea are not currently thought to contain species of medical or veterinary importance, they will not be considered further. Blue–green algae do not cause infection, but certain species produce potent peptide toxins that may affect persons or animals ingesting polluted water.
The viruses are the smallest of the infective agents; they have a relatively simple structure that is not comparable with that of a cell, and their mode of reproduction is fundamentally different from that of cellular organisms. Even simpler are viroids , protein-free fragments of single-stranded circular RNA that cause disease in plants. Another class of infectious particles are prions , the causative agents of fatal neurodegenerative disorders in animals and man. These are postulated to be naturally occurring host cell membrane glycoproteins that undergo conformative changes to an infectious isoform (see Ch. 60).
Taxonomy consists of three components: classification, nomenclature and identification . Classification allows the orderly grouping of micro-organisms, whereas nomenclature concerns the naming of these organisms and requires agreement so that the same name is used unambiguously by everyone. Changes in nomenclature may give rise to confusion and are subject to internationally agreed rules. In clinical practice, microbiologists are generally concerned with identification – the correct naming of isolates according to agreed systems of classification. These components, together with taxonomy, make up the overarching discipline of systematics , which is concerned with evolution, genetics and speciation of organisms, and is commonly referred to as phylogenetics .
Protozoa, fungi and helminths are classified and named according to the standard rules of classification and nomenclature that have been developed following the pioneering work of the eighteenth century Swedish botanist Linnaeus (Carl von Linné). Large subdivisions (class, order, family, etc.) are finally classified into individual species designated by a Latin binomial, the first term of which is the genus , e.g. Plasmodium (genus) falciparum (species). Occasionally it is useful to recognize a biological variant with particular properties: thus, Trypanosoma (genus) brucei (species) gambiense (variant) differs from the variant T. brucei brucei in being pathogenic for man.
Bacteria are similarly classified, but bacterial diversity encompasses more variety than all the rest of cellular life put together and the natural capacity of bacteria for genetic and phenotypic variation and adaptation make rigid classifications difficult. To date, identification has predominantly been performed by the use of keys that allow the organization of bacterial traits based on growth or activity in systems that test their biochemical properties. Some tests are definitive of a genus or species, for example the universal production of catalase enzyme and cytochrome c, respectively, by Staphylococcus spp. and Pseudomonas aeruginosa . Other characters may be unique to individual species and serve to differentiate them from organisms with closely similar biochemical activity profiles. Some bacteria do not grow in the laboratory (leprosy bacillus, treponemes), and identification by genetic methods may be necessary. As the technologies for genetic analysis become more readily applicable in clinical labs, so they and other rapid analytical methods, such as those based on mass spectrometry, are coming to replace the traditional biochemical methods to achieve identification. The taxonomic ranks used in the classification of bacteria are (example in parentheses):
- • Kingdom (Prokaryotae)
- • Division (Gracilicutes)
- • Class (Betaproteobacteria)
- • Order (Burkholderiales)
- • Family (Burkholderiaceae)
- • Genus ( Burkholderia )
- • Species ( Burkholderia cepacia ).
Some genera, such as Acinetobacter , have been subdivided into a number of genomic species by DNA homology analysis. Some are named and others are referred to only by a number. Many of the genomic species cannot be differentiated with accuracy by phenotypic tests. Another subgenus grouping in current usage recognizes species complexes, which are differentiated into genomovars by polyphasic taxonomic methods. A good example of this is the B. cepacia complex of organisms, which includes a very diverse group of organisms ranging from strict plant to human pathogens.
At present no standard classification of bacteria is universally accepted and applied, although Bergey's Manual of Determinative Bacteriology is widely used as an authoritative source. Bacterial nomenclature is governed by an international code prepared by the International Committee on Systematic Bacteriology and published as Approved Lists of Bacterial Names in the International Journal of Systematic and Evolutionary Microbiology ; most new species are also first described in this journal, and a species is considered to be validly published only if it appears on a validation list in this journal.
The International Committee on Taxonomy of Viruses (ICTV) classifies viruses and publishes its reports in the journal Archives of Virology . Latin names are used wherever possible for the ranks family, subfamily and genus, but at present there are no formal categories higher than family and binomial nomenclature is not used for species. Viruses do not lend themselves easily to classification according to Linnaean principles, and vernacular names still have wide usage among medical virologists. Readers are referred to the standard work on virus taxonomy Classification and Nomenclature of Viruses and the ICTV database website.
Methods of classification
Adansonian or numerical classification.
In most systems of bacterial classification, the major groups are distinguished by fundamental characters such as cell shape, Gram-stain reaction and spore formation; genera and species are usually distinguished by properties such as fermentation reactions, nutritional requirements and pathogenicity. The relative ‘importance’ of different characters in defining major and minor groupings is often purely arbitrary. The uncertainties of arbitrary choices are avoided in the Adansonian system of taxonomy. This system determines the degrees of relationship between strains by a statistical coefficient that takes account of the widest range of characters, all of which are considered of equal weight. It is clear, of course, that some characters, for instance cell shape or Gram-stain reaction, represent a much wider and permanent genetic commitment than other characters which, being dependent on only one or a few genes, may be unstable. For this reason, the Adansonian method is most useful for the classification of strains within a larger grouping that shares major characters.
By scoring a large number of phenotypic characters it is possible to estimate a similarity coefficient when shared positive characters are considered or a matching coefficient when both negative and positive shared characters (matches) are taken into account. This numerical taxonomy is best performed on a computer that can calculate degrees of similarity for a group of different organisms; these data are displayed as a similarity matrix or a dendrogram tree ( Fig. 3.1 ) (see also Owen 2004 in Recommended reading list).

Hierarchical taxonomic tree ( dendrogram ) prepared from similarity matrix data. The dashed lines X and Y indicate levels of similarity at which separation into genera and species might be possible.
DNA composition
The hydrogen bonding between guanine and cytosine (G–C) base pairs in DNA is stronger than that between adenine and thymine (A–T). Thus, the melting or denaturation temperature of DNA (at which the two strands separate) is determined primarily by the G + C content. At the melting temperature, the separation of the strands brings about a marked change in the light absorption characteristics at a wavelength of 260 nm, and this is readily detected by spectrophotometry. There is a very wide range in the G + C component of bacterial DNA, varying from about 25–80% mol in different genera. However, for any one species, the G + C content is relatively fixed, or falls within a very narrow range, and this provides a basis for classification.
DNA homology
Another approach to classification is to arrange individual organisms into groups on the basis of the homology of their DNA base sequences. This exploits the fact that double strands re-form (anneal) from separated strands during controlled cooling of a heated preparation of DNA. This process can be readily demonstrated with suitably heated homologous DNA extracted from a single species, but it can also occur with DNA from two related species, so that hybrid pairs of DNA strands are produced. These hybrid pairings occur with high frequency between complementary regions of DNA, and the degree of hybridization can be assessed if labelled DNA preparations are used. Binding studies with messenger RNA (mRNA) can also give information to complement these observations, which provide genetic evidence of relatedness among bacteria. Organisms with different G + C ratios are unlikely to show significant DNA homology. However, organisms with the same, or close, G + C ratios do not necessarily show homology. A novel real-time polymerase chain reaction (PCR) (see pp. 31 and 79–80) has been described for estimation of G + C content.
Ribosomal RNA sequencing
The structure of ribosomal RNA (rRNA) appears to have been highly conserved during the course of evolution, and close similarities in nucleotide sequences reflect phylogenetic relationships. Advances in technology have made nucleotide sequencing relatively simple, and the rDNA sequences (and other genes) of most medically important bacterial species are available from a number of internet sites. Far fewer full-length gene sequences are known for 23S rRNA than for 16S rRNA and 16S–23S internal transcribed sequences. In practice the DNA of the test organism is extracted and amplified by PCR using universal primers. The DNA sequence of the product is determined and the sequence compared against databases to find the closest fit. It is generally accepted that sequence similarity between 0.5% and 1.0% (with the type species) is required for the identification of an unknown organism, and less than 97% similarity is a common cut-off point for the differentiation of species. However, different species of Mycobacterium may exhibit more than 97% similarity in rDNA sequence. Commercial systems are available for bacterial species identification (MicroSeq; Applied Biosystems, Foster City, California, USA).
Nucleotide sequence variation in ribosomal genes is sometimes insufficient to discriminate between closely related species. Other candidate genes have been explored but the recA gene, which encodes a protein essential for repair and recombination of DNA, appears to be one of the best suited for phylogenetic analysis in that it defines evolutionary trees consistent with those observed for rRNA genes. Additional housekeeping genes used for phylogenetic studies include rpoB (RNA polymerase), groEL (heat shock protein) and gyrB (DNA gyrase), among others.
Classification in clinical practice
The identification of micro-organisms in routine practice requires a pragmatic approach to taxonomy. The primary purpose of the names used is to communicate amongst the clinical and public health teams so that appropriate management of individuals and groups can be implemented. Unfortunately, as we learn more about the interrelatedness of microbes, largely through genome sequence data, certain reallocation and renaming becomes necessary in order to sustain a coherent relationship with our basic understanding. Table 3.1 outlines a simple, but practical, classification scheme in which organisms are grouped according to a few shared characteristics; as far as possible these have been reconciled with the relatively recently established phylogenetic nomenclature. Within these groups, organisms may be further identified, sometimes to species level, by a few supplementary tests. Protozoa, helminths and fungi can often be definitively identified on morphological criteria alone (see appropriate chapters).
Simplified classification of some cellular micro-organisms of medical importance
These are non-photosynthetic unicellular organisms with protoplasm clearly differentiated into nucleus and cytoplasm. They are relatively large, with transverse diameters mainly in the range of 2–100 µµm. Their surface membranes vary in complexity and rigidity from a thin, flexible membrane in amoebae, which allows major changes in cell shape and the protrusion of pseudopodia for the purposes of locomotion and ingestion, to a relatively stiff pellicle in ciliate protozoa, preserving a characteristic cell shape. Most free-living and some parasitic species capture, ingest and digest internally solid particles of food material; many protozoa, for instance, feed on bacteria. Protozoa, therefore, are generally regarded as the lowest forms of animal life, although certain flagellate protozoa are closely related in their morphology and mode of development to photosynthetic flagellate algae in the plant kingdom. Protozoa reproduce asexually by binary fission or multiple fission ( schizogony ), and some also by a sexual mechanism ( sporogony ). The most important groups of medical protozoa are the sporozoa (malaria parasites, etc.), amoebae and flagellates (see Ch. 62).
These are non-photosynthetic organisms that possess relatively rigid cell walls. They may be saprophytic or parasitic, and take in soluble nutrients by diffusion through their cell surfaces.
Moulds grow as branching filaments ( hyphae ), usually 2–10 µm in width, which interlace to form a meshwork ( mycelium ). The hyphae are coenocytic (i.e. have a continuous multinucleate protoplasm), being either non-septate or septate with a central pore in each cross-wall. Moulds reproduce by the formation of various kinds of sexual and asexual spores that develop from the vegetative (feeding) mycelium, or from an aerial mycelium that effects their air-borne dissemination (see Ch. 61).
Yeasts are ovoid or spherical cells that reproduce asexually by budding and also, in many cases, sexually, with the formation of sexual spores. They do not form a mycelium, although the intermediate yeast-like fungi form a pseudo-mycelium consisting of chains of elongated cells. The dimorphic fungi produce a vegetative mycelium in artificial culture, but are yeast-like in infected lesions. The higher fungi of the class Basidiomycetes (mushrooms), which produce large fruiting structures for aerial dissemination of spores, are not infectious for human beings or animals, although some species are poisonous.
The main groups of bacteria are mostly distinguished by microscopic observation of their morphology and staining reactions. The Gram-staining procedure, which reflects fundamental differences in cell wall structure, separates most bacteria into two great divisions: Gram-positive bacteria and Gram-negative bacteria (see Ch. 2). Where bacteria can be Gram stained the cell shape and clustering are of practical value in identification. However, at this point the relationship of the older, largely phenotypic, classification system to the newer DNA sequence-based phylogenetic classification, which reflects the evolutionary relationships between groups, begins to break down and anomalies are apparent. Nonetheless, practical identification schemes used in clinical labs rely heavily on recognizing whether bacteria are Gram positive or negative, bacillary or coccal in shape, able to grow aerobically or anaerobically (see Ch. 4) and whether they form spores.
The major phylogenetic groupings of medically significant bacteria are:
- • Actinobacteria , these are also recognised as Gram-positve bacteria with a high G + C content, many of which are capable of filamentous growth with true branching and which may produce a type of mycelium. Many members of this group do not stain well with the Gram method and, importantly, they include the mycobacteria which can be recognized with acid fast stains (see Ch. 2).
- • Firmicutes , low G + C Gram-positive bacteria including bacilli, some of which are spore formers and cocci; the group includes most of the medically significant Gram-positives. Two groups that have presumably lost their Gram-positivity, Veillonella and Mycoplasma are included here.
- • Proteobacteria , a very large of group of Gram-negative bacteria (bacilli and cocci) with five subdivisions (alpha, beta, gamma, delta and epsilon).
- • Bacteroidetes , Gram-negative anaerobes.
- • Spirochaetes , possessing cells with a tight spiral shape and an internal flagellum.
- • C hlamydiae , which are strict intracellular parasites.
Actinobacteria
These include antibiotic producing bacteria and a small number of highly significant pathogens.
- • Actinomyces. Gram-positive, non-acid-fast, tend to fragment into short coccal and bacillary forms and not to form conidia; anaerobic (e.g. Actinomyces israelii ).
- • Streptomyces. Vegetative mycelium does not fragment into short forms; conidia form in chains from aerial hyphae (e.g. Streptomyces griseus ).
- • Mycobacterium. Acid-fast; Gram-positive, but does not readily stain by the Gram method; usually bacillary, rarely branching; make mycolic acids (e.g. Mycobacterium tuberculosis ).
- • Nocardia . Similar to Actinomyces , but aerobic and mostly acid-fast; make mycolic acids (e.g. Nocardia asteroides ).
- • Corynebacterium. Pleiomorphic (variably shaped) Gram-positive bacilli; make mycolic acids (e.g. Corynebacterium diphtheriae ).
- • Streptococcus . Gram-positive cocci, mainly adherent in chains due to successive cell divisions occurring in the same axis (e.g. Streptococcus pyogenes ); sometimes predominantly diplococcal (e.g. Streptococcus pneumoniae ).
- • Staphylococcus . Gram-positive cocci, mainly adherent in irregular clusters due to successive divisions occurring irregularly in different planes (e.g. Staph. aureus ).
- • Mycoplasma and ureoplasma. Pleiomorphic cocci that do not make peptidoglycan.
- • Veillonella. Gram-negative; generally very small cocci arranged mainly in clusters and pairs; anaerobic (e.g. Veillonella parvula ).
- • Gram-positive spore-forming bacilli. The key medically relevant genera here the genera Bacillus and Clostridium (anaerobic). They are Gram positive, but liable to become Gram negative in ageing cultures. The size, shape and position of the spore may assist recognition of the species, for example the bulging, spherical, terminal spore ('drumstick’ form) of Clostridium tetani .
- • Gram-positive non-sporing bacilli. These include several genera. Erysipelothrix and Lactobacillus are distinguished by a tendency to grow in chains and filaments, and Listeria by flagella that confer motility.
Proteobacteria
Gram-negative bacteria.
- • Alphaproteobacteria. Include the cell dependent Rickettsia group, the facultatively intracellular Brucella group and the Bartonella group.
- • Betaproteobacteria. Neisseria – cocci, mainly adherent in pairs and slightly elongated at right angles to axis of pairs (e.g. Neisseria meningitidis ). Burkholderia – bacilli ( Burkholderia pseudomallei).
- • Gammaproteobacteria . Bacilli including the enterobacteria ( Escherichia coli) , and the genera Pseudomonas , Legionella and the curved vibrios (e.g. Vibrio cholera ).
- • Deltaproteobacteria . No medically significant bacteria.
- • Epsilonproteobacteria . Curved and loosely spiral bacilli including the genera Helicobacter and Campylobacter .
Bacteroidetes
Gram-negative non-sporing anaerobic bacilli.
- • Bacteroides , Prevotella and Porphyromonas are major genera.
Spirochaetales
These organisms differ from the other groups in being slender flexuous spiral filaments that, unlike the spirilla, are motile without possession of flagella. The staining reaction, when demonstrable, is Gram negative. The different varieties are recognized by their size, shape, waveform and refractility, observed in the natural state in unstained wet films by dark-ground microscopy. Genera of medical importance include Borrelia , Treponema and Leptospira .
Chlamydiae have a complex intracellular cycle (e.g. Chlamydia trachomatis ).
Viruses usually consist of little more than a strand of DNA or RNA enclosed in a simple protein shell known as a capsid . Sometimes the complete nucleocapsid may be enclosed in a lipoprotein envelope derived largely from the host cell. Viruses are capable of growing only within the living cells of an appropriate animal, plant or bacterial host; none can grow in an inanimate nutrient medium. The viruses that infect and parasitize bacteria are termed bacteriophages or phages . A simple classification of the viruses that are involved in human disease is shown in Table 3.2 .
Principal types of virus causing human disease
SARS, severe acute respiratory syndrome.
Identification of micro-organisms
Precise or definitive identification of bacteria is time consuming, contentious and best carried out in specialized reference centres. For most clinical purposes, clear, rapid guidance on the likely cause of an infection is required and, consequently, microbiologists usually rely on a few simple procedures, notably microscopy and culture, backed up, when necessary, by a few supplementary tests to achieve a presumptive identification. Microscopy is the most rapid test of all, but culture inevitably takes at least 24 hours, sometimes longer. More rapid tests are constantly being sought, and antigen detection methods and genetic detection methods are now well established (see below).
Most specimens for bacteriological examination, whether from human beings, animals or the environment, contain mixtures of bacteria, and it is essential to obtain pure cultures of individual isolates before embarking on identification. Non-cultural methods, such as antigen or nucleic acid based detection, do not have this disadvantage; however, they do have the potential limitation of being highly specific so that the investigator must know beforehand what it is necessary to look for.
Morphology and staining reactions of individual organisms generally serve as preliminary criteria to place an unknown species in its appropriate biological group. A Gram-stain smear suffices to show the Gram reaction, size, shape and grouping of the bacteria, and the arrangement of any endospores. An unstained wet film may be examined with dark-ground illumination in the microscope to observe the morphology of delicate spirochaetes; an unstained wet film, or ‘hanging-drop’, preparation is examined with ordinary bright-field illumination for observation of motility. Capsules surrounding bacterial cells are demonstrated by ‘negative staining’ with India ink; the capsules remain unstained against the background of ink particles. To identify mycobacteria, or other acid-fast organisms, a preparation is stained by the Ziehl–Neelsen method or one of its modifications (see p. 16). The microscopic characters of certain organisms in pathological specimens may be sufficient for presumptive identification, for example tubercle bacilli in sputum, or T. pallidum in exudate from a chancre. However, many bacteria share similar morphological features, and further tests must be applied to differentiate them.
Cultural characteristics
The appearance of colonial growth on the surface of a solid medium, such as nutrient agar, is often very characteristic. Attention is paid to the diameter of the colonies, their outline, their elevation, their translucency (clear, translucent or opaque) and colour. Changes brought about in the medium (e.g. haemolysis in a blood agar medium) may also be significant. The range of conditions that support growth is characteristic of particular organisms. The ability or inability of the organism to grow in the presence (aerobe) or absence (anaerobe) of oxygen, in a reduced oxygen atmosphere (micro-aerophile) or in the presence of carbon dioxide, or on media containing selective inhibitory factors (e.g. bile salt, specific antimicrobial agents, or low or high pH) may also be of diagnostic significance (see Table 4.1).
Biochemical reactions
Species that cannot be distinguished by morphology and cultural characters may exhibit metabolic differences that can be exploited. It is usual to test the ability of the organism to produce acidic and gaseous end-products when presented with individual carbohydrates (glucose, lactose, sucrose, mannitol, etc.) as the sole carbon source. Other tests determine whether the bacterium produces particular end-products (e.g. indole or hydrogen sulphide) when grown in suitable culture media, and whether it possesses certain enzyme activities, such as oxidase, catalase, urease, gelatinase or lecithinase. Traditionally, such tests have been performed selectively and individually according to the recommendations of standard guides, such as the invaluable Cowan and Steel's Manual for the Identification of Medical Bacteria . However, today most diagnostic laboratories use commercially prepared microgalleries of identification tests which, though expensive, combine simplicity and accuracy. Test kits are now available for a number of different groups of organisms, including enterobacteria, staphylococci, streptococci and anaerobes. Other kits facilitate the testing of carbon source utilization, the assimilation of specific substrates and the enzymes produced by an organism.
On occasion, more elaborate procedures may be used for the analysis of metabolic products or whole-cell fatty acids. Indeed, a fully automated, fatty acid-based identification system, which combines high-resolution gas chromatography and pattern recognition software, is widely used to identify a variety of aerobic and anaerobic bacterial species. New profiles are added to a computerized database, thus increasing the sensitivity of the system. Mass spectrometric methods show promise for rapid identification, particularly matrix-assisted laser desorption ionization time-of-flight (MALDI-TOF) mass spectrometry. This offers the analysis of whole bacterial cultures for unique mass spectra from charged macromolecules by rapid, high-throughput testing with a rapidly growing database.
Indirect identification methods
Gene-targeted analyses.
These are fragments of DNA that recognize complementary sequences within micro-organisms (see p. 79). Binding is detected by tagging the DNA with a radioactive label, or with a reagent that can be developed to give a colour reaction, such as biotin. By selecting DNA fragments specific for features characteristic of individual organisms, gene probes can be tailored to the rapid identification of individual species in clinical material. The disadvantages of this approach are that the organism itself may be dead and a viable isolate is not made available for subsequent tests of susceptibility to antimicrobial agents, toxin production or epidemiological investigation.
Culture and preliminary identification of bacteria in the laboratory is time consuming and relatively labour intensive. Bacteria may also be uncultivable, slow growing, or fastidious in nutrient requirements. Nucleic acid techniques for the detection and identification of bacteria have evolved against this background; today numerous commercially available systems have been developed and many are in use in diagnostic laboratories. The technologies fall into three basic groups:
- 1 Target amplification by PCR, transcription-mediated amplification, nucleic acid sequence-based amplification, etc.
- 2 Probe amplification using ligase chain reaction or Q-beta replicase.
- 3 Signal amplification, as in branched DNA assay.
It is possible to detect the presence of an increasing number of species either by PCR of universal or specific gene targets or by hybridization with specific probes. As with conventional methods, nucleic acid technology has its limitations, the most frequent being contamination of a sample by post-amplification products. Other factors include operator skill, primer design, stringency of assay, presence of inhibitory compounds in the specimen and the ubiquity of the organism sought. The latter is fundamental to the interpretation of results as many bacterial pathogens occur naturally as commensals in certain body sites. The new technologies do, however, offer a considerable advantage over phenotypic methods in terms of sensitivity, and many optimized PCR systems claim to be able to detect as few as two to ten bacteria per millilitre of specimen, which is far below the threshold of conventional culture.
Nucleic acid assays for antimicrobial resistance genes are also in use and development. A recently described innovation may have the potential to detect, identify the species, subtype the organism and identify resistance genes on microscope slide smears. The technique uses peptide nucleic acid (PNA, pseudopeptides) with DNA binding capacity. The PNA molecules have a polyamide backbone instead of the sugar phosphate of DNA and RNA, and nucleotide bases attached to this backbone are able to hybridize by specific base pairing with complementary DNA or RNA sequences. PCR amplification of target genes on conventionally stained microscope smears is also possible, and this accelerates the prospect of very rapid and sensitive test methods limited only by the specificity of the primer for the target.
The availability of numerous prokaryotic genomic sequences has enabled the development of high-density oligonucleotide arrays which consist of many thousands of different probes. These arrays are constructed by in-situ oligonucleotide synthesis on a glass support by photolithography or other methods. Hybridization of the prelabelled target nucleic acids to the bound probe is detected directly with fluorescein or radioactive ligands, or indirectly using enzyme conjugates. Areas of application for high-density arrays include DNA sequencing, strain genotyping, identifying gene functions, location of resistance genes, changes in mRNA expression and phylogenetic relatedness. Arrays with selected gene targets have recently been developed in an Eppendorf tube format. The chip is embedded in the bottom of the tube and carries optimized sets of oligonucleotide probes specific for certain organisms or for antimicrobial resistance genes or virulence factors. In this way chips can be customized for individual bacteria or groups of bacteria. All stages of the assay – sample preparation from agar-grown colonies, PCR amplification, hybridization, conjugation with reporter molecule and detection by automated image recording – are carried out in a single tube within 6–8 hours.
A widely used development is that of real-time PCR, which combines sample amplification with a means for detection of the specific product by fluorescence so that both steps take place conveniently in a single reaction tube. The system has significant advantages over conventional PCR in terms of rapidity, simplicity and number of manual procedures; contamination is effectively eliminated by the tube being closed after amplification. The DNA product may be detected with a fluorescent dye, or increased specificity obtained with hybridization with fluorescence-labelled sequence-specific oligonucleotide probes. Quantification of the target DNA is also possible with this system, allowing estimation of viral or bacterial numbers in a specimen (see p. 79–80).
Fluorescence in-situ hybridization (FISH) directed to the multiple copies of 16S ribosomal RNA has also been used to detect bacteria directly in clinical specimens. This technique utilizes probes specific for target organisms in the sample without the need for culture and allows quantification of the cell count and morphology. Sensitivity and specificity vary according to the probe used, but if the probe is optimized it can rival conventional culture methods.
Antibody reactions
Species and types of micro-organism can often be identified by specific serological reactions. These depend on the fact that the serum of an animal immunized against a micro-organism contains antibodies specific for the homologous species or type that react in a characteristic manner (e.g. agglutination or precipitation) with the particular micro-organism. Such simple in-vitro tests have been used for many years in microbiology, notably in the formal identification of presumptive isolates of pathogens (e.g. salmonellae) from clinical material. The specificity and range of antibody tests have been greatly improved by the availability of highly specific monoclonal antibodies . These are produced by the hybridoma technique in which individual antibody-producing spleen cells are fused with ‘immortal’ tumour cells in vitro. The progeny of these hybrid cells produce only the type of antibody appropriate to the spleen cell precursor (see p. 125).
Latex agglutination
By adsorbing specific antibody to inert latex particles, a visible agglutination reaction can be induced in the presence of homologous antigen. This principle can be applied in reverse to detect serum antibodies. Latex-based kits are widely used for serological grouping of organisms and detection of toxins produced by bacteria during growth.
Enzyme-linked immunosorbent assay
In enzyme-linked immunosorbent assay (ELISA), a specific antibody is attached to the surface of a plastic well and material containing the test antigen is added. After washing, the presence of the antigen is detected by addition of more of the specific antibody, this time labelled with an enzyme that can initiate a colour reaction when provided with the appropriate substrate. The intensity of the colour change is related to the amount of antigen bound. The ELISA method may also be used in the reverse manner for the quantitative detection of antibodies, by adsorbing purified antigen to the well before adding test serum; in this case the enzyme-linked system used to detect the antigen–antibody reaction is a labelled anti-human globulin. In immunoglobulin (Ig) M antibody capture ELISA (MAC-ELISA), widely used in virological diagnosis for the detection of IgM, anti-human µ-chain antibody (usually raised in goats) is bound to the well. The test serum is added, and any IgM binds to the capture reagent; after washing, purified antigen (e.g. rubella antigen) is added, and this can be detected with an appropriate labelled antibody.
Haemagglutination and haemadsorption
Certain viruses, notably the influenza viruses, have the property of attaching to specific receptors on the surface of appropriate red blood cells. In this manner the virus particles act as bridges linking the red cells in visible clumps. In tissue culture, such haemagglutinins may appear on the surface of cells infected with a virus. If red cells are added to the tissue culture, they adhere to the surface of infected cells, a phenomenon known as haemadsorption . Red blood cells can also be coated with specific antibody so that they agglutinate in the presence of the homologous virus particle in a manner similar to that described for latex agglutination above.
Fluorescence microscopy and immunofluorescence
When certain dyes are exposed to ultraviolet light, they absorb energy and emit visible light; that is, they fluoresce. Tissues or organisms stained with such a dye and examined with ultraviolet light in a specially adapted microscope are seen as fluorescent objects; for example, auramine can be used in this way to stain Mycobacterium tuberculosis . Antibody molecules can be labelled by conjugation with a fluorochrome dye such as fluorescein isothiocyanate (which fluoresces green) or rhodamine (orange–red). When fluorescent antibody is allowed to react with homologous antigen exposed at a cell surface, this direct immunofluorescence procedure affords a highly sensitive method for the identification of the particular antigen. For this procedure it is necessary to have a specific antibody conjugate for each antigen; however, unconjugated antibody can be used and the reaction then detected by the addition of an antiglobulin conjugate, which will react with any antibody from the species in which the antibody was raised.
Immuno-polymerase chain reaction
This technique arose out of the fusion of antibody technology with PCR methods with the aim of enhancing the capability of antigen detection systems. In immuno-PCR, a linker molecule with bispecific binding affinity for DNA and antibodies is used to attach a DNA molecule (marker) to an antigen–antibody complex. This produces a specific antigen–antibody–DNA conjugate. The attached DNA marker can be amplified by PCR with appropriate primers, and the presence of amplification products shows that the marker DNA is attached to antigen–antibody complexes, indicating the presence of antigen. The enhanced sensitivity of immuno-PCR achieved over ELISA is reported to be in excess of 10 5 , theoretically allowing as few as 580 antigen molecules (9.6 × 10 −22 moles) to be detected.
Typing of bacteria
Different bacterial species often exhibit different population structures. Some species are characterized by highly diverse populations at one extreme and closely similar members at the other. The frequency of recombination of chromosomal genes (see Ch. 6) is considered the major determinant of a population structure of a given species, and this frequency ranges from absent to low to very high. Highly recombining populations are termed panmictic , in contrast to clonal populations where recombination is infrequent ( Table 3.3 ). Species such as Neisseria gonorrhoeae and Haemophilus influenzae are naturally transformable, that is, they are able to take up DNA (foreign and native) from their environment, and their populations are characterized by a high frequency of recombination, segregation of alleles and relatively low mutation. In clonal populations such as Salmonella enterica, recombination is rare and there is non-random association of alleles in a background of limited genetic exchange. Mutations occur as a result of natural and selective pressures, but these are not sufficient to disrupt the clonal lineage and daughter cells continue to resemble the ancestral parent. Bacterial clones are therefore not identical to their parents but display a number of characteristics in common with their ancestors. Many species are characterized by considerable genetic diversity but with clonal expansion of a subpopulation. Some of these clones may be transient, although others may persist and spread nationally and globally.
Panmictic versus clonal populations
By typing we identify a recognizable subdivision of a species that serves as a reference marker against which other isolates of the same species can be compared. A population of bacteria presumed to descend from a single bacterium, as found in a natural habitat, in primary cultures from the habitat, and in subcultures from the primary cultures, is called a strain. Each primary culture from a natural source is called an isolate . The distinction between strains and isolates may be important; for example, cultures of typhoid bacilli isolated from ten different patients should be regarded simply as ten different isolates unless epidemiological or other evidence indicates that the patients have been infected from a common source with the same strain . The ability to discriminate between similar strains may be of great epidemiological value in tracing sources or modes of spread of infection in a community or hospital ward, and various typing methods have been devised. Strains may be distinguishable only in minor characters and it is usually simpler to establish differences between isolates from a common source than unequivocally to prove their identity. Demonstration of an identical response by a single reproducible typing method is not proof that two strains are the same. However, the confidence with which similarity can be inferred is greatly increased if more than one typing method is used.
Typing may inform different levels of epidemiological investigation, ranging from micro-epidemiology (local investigation), macro-epidemiology (regional, national, international) to population structure analysis (evolution of strains and global patterns of spread). The data derived may assist in the control of infection by excluding sources, identifying carriers and establishing the prevalence of individual strains. Common reasons for microbial typing are to identify common or point sources, discriminate between mixed strain infections, distinguish re-infection from relapse, and occasionally to identify a type and disease association (e.g. Escherichia coli O157 and haemolytic uraemic syndrome, skin and throat types of group A Str. pyogenes , etc.).
Typing methods should be reproducible both in the laboratory and clinically. The former is easily established by repeated tests on a sample of experimental strains, but should also be established in vivo by examining multiple pairs of isolates from single sources to determine the stability of the strain characteristics probed by the typing method used. A typing method should also discriminate adequately and clearly between different populations and be comprehensive, that is, assign most populations to a type. The typing data should be in a format that is easily assimilated into databases and should be able to be incorporated into the national picture to inform other workers in the field. Very few, if any, single typing methods will meet these criteria and hence there is a need to utilize different methods, preferably directed at unlinked targets and always in the context of an epidemiological investigation.
Biochemical test reactions that are not universally positive or negative within a species may define biotypes of the species, and these may be efficient strain markers. In practice biotyping is often less discriminatory than other strain typing methods and may be unstable because of loss of the property. Differences among strains may also be detected by variations in sensitivity to fixed concentrations of chemicals such as heavy metals, a process known as resistotyping . The nutritional requirements of the isolate (amino acids) for growth may also be used to define the auxotype of an isolate.
Many surface structures of bacteria (lipopolysaccharide and outer membrane, flagella, capsule, etc.) are antigenic, and antibodies raised against them can be used to group isolates into defined serotypes . Some species are characterized by numerous antigenic types and serotyping for these species is highly discriminatory, whereas for others conservation of antigen epitopes renders serotyping of little value for epidemiological purposes. Members of the species Salmonella enterica are defined by their somatic and flagellar serotypes (see Ch. 24). Capsular antigens may be associated with pathogenicity of the organism, and many vaccines protect the individual against infection by stimulating antibodies to capsular antigen epitopes. Agglutination of bacterial suspensions with rabbit antibodies is the most commonly used method for typing, but other techniques such as precipitation in agar gels, ELISA and capsular swelling ( Fig. 3.2 ) may be used.

Capsular swelling reaction of Klebsiella pneumoniae . Antibody adsorbed to capsule alters the refractive index, allowing visualization of the capsule around the cell within.
Phage typing
Bacteria often show differential susceptibility to lysis by certain bacteriophages. The phage adsorbs to a specific receptor on the bacterial surface and injects its DNA into the host. Phage DNA may become stably integrated into the bacterial chromosome and this state is referred to as lysogeny ; phages capable of this are called temperate phages . In lysogeny, a small proportion of host cells express the phage genes, and some cell lysis and liberation of phage progeny occurs. Alternatively, the phage DNA may enter a replicative cycle, leading to the death of the host and the production of new phage particles. These lytic or virulent wild phages lyse the bacterium at the end of the replicative cycle and release a large number of daughter phage particles that infect neighbouring cells.
This process leads to visible inhibition of the growth of the host cells ( Fig. 3.3 ). The phage type of the culture is identified according to the pattern of susceptibility to a set of lytic and/or temperate phages. Lytic phages may be readily recovered from sewage, waste and river water, and temperate phages may be released from a lysogenic strain by induction with ultraviolet radiation or chemical mutagens.

Phage-mediated lysis of red pigmented strain of Serratia marcescens .
The critical factors governing the interpretation of phage typing results are discrimination and reproducibility. If the system is both highly discriminatory and reproducible, any differences in lysis patterns of isolates will indicate that they represent different strains. Schemes that utilize adapted phages (a single phage propagated in different strains) that are specific for a particular receptor site such as the Vi polysaccharide of Salmonella enterica serotype Typhi are relatively reproducible, and minor differences are significant and reproducible. On the other hand, a phage set comprising random unrelated phages may adhere to a number of different receptor sites and have different biological properties, and so is unlikely to be highly reproducible but may be adequately discriminating. This lack of stability results in the definition of broad phage groups rather than defined types, and is the case for Staph. aureus where at least two strong lytic reactions have to be present in the patterns of isolates before they can be termed distinct strains.
Bacteriocin typing
Bacteriocins are naturally occurring antibacterial substances, elaborated by most bacterial species, that are active mainly against strains of the same genus as the producer strain. Bacteriocin typing may define the spectrum of bacteriocins produced by field strains, or the sensitivity of these strains to bacteriocins of a standard panel of strains. Patterns of production or susceptibility to bacteriocins allow the division of species into bacteriocin types.
Protein typing
Bacteria manufacture thousands of proteins that can be visualized by electrophoresis in acrylamide gels in the presence of a strong detergent. The proteins separate according to molecular size and, after staining with a dye, the pattern of bands from each isolate can be compared. This system has been used successfully to type many bacterial and fungal species, but lacks reproducibility. Investigation of microbial populations by gel electrophoresis of metabolic enzymes, which can then be detected by specific substrates, has also in the past been applied widely for clonal analysis within species.
Restriction endonuclease typing
Restriction endonucleases are a family of enzymes that each cut DNA at a specific sequence recognition site, which may be rare or frequent in the DNA of the species being examined. The frequency with which an enzyme cuts in a particular species is dependent on the oligonucleotide sequence, the frequency of the restriction site, and the percentage G + C content of the species. For example, the recognition site of enzyme Sma I is 5′-CCC↓GGG-3′ (↓ site of cleavage) and this cuts infrequently in the AT-rich genome of Staph. aureus , whereas enzyme Xba I (5′-T↓CTAGA-3′) is a rare cutter in most Gram-negative species with a high GC content. Both plasmid and chromosomal DNA can be analysed by this means. Frequent-cutting endonucleases generate numerous small fragments that can be resolved by conventional electrophoresis in agarose gel and detected by staining with a dye. The resolution of conventional agarose gel electrophoresis does not exceed 20 kb and optimal separation in standard length gels is achieved between 1 and 15 kb.
The large DNA fragments produced by infrequent-cutting enzymes need to be separated in special electrical fields with a pulsed current ( pulsed-field gel electrophoresis ; PFGE). In this technique, bacteria are encased in an agarose plug (to minimize shearing of DNA) and the cells are digested with proteinase K enzyme before the DNA is digested with the enzyme. By introducing a pulse or change in the direction of the electric field, fragments as large as 10 Mb can be separated. The time taken by fragments to reorient to the alternate electric field is proportional to their molecular size and where they migrate in the electric field. The most widely used apparatus is the contour-clamped homogeneous electric field (CHEF), which has 24 electrodes arranged in a hexagonal array. Run times are often of the order of 30–40 hours, but shorter, more rapid, protocols have been described. A number of factors influence the quality of results, including DNA quality and concentration, agarose concentration, voltage and pulse times, and buffer strength and temperature.
Interpretation of PFGE profiles can be problematic. For some species the criteria of Tenover (see recommended reading list) can be applied to establish the significance of differences in banding profiles of strains. As a rule of thumb, isolates from an incident under investigation that show no difference in profiles can be considered indistinguishable, those with one to three band differences as closely related, four to six bands as possibly related, and seven or more band differences as indicating distinct strains. However, this rule should be applied with a degree of caution as some species (e.g. Enterococcus faecium ) can exhibit significant variation (six to ten band differences) apparently within members of the same clone. A number of computer-assisted analysis packages are available that calculate coefficients of similarity between strains and represent these as dendrograms ( Fig. 3.4 ). Two commonly employed coefficients, the Jaccard and Dice, use the number of concordant bands in profiles and the total number of possible band positions to calculate the percentage similarity between the isolates. The Pearson coefficient gives the advantage that specific band positions do not have to be defined. A cut-off point of 85% similarity is often used but, as for the band difference rule, this should be set by experiment with related and unrelated strain sets.

Pulsed-field gel electrophoresis profiles of Xbal digests of DNA of Ps. aeruginosa isolates. Dendrogram shows relative percentage similarity of profiles calculated with Pearson's coefficient.
Gene probe typing
DNA probes (see above) for strain typing consist of cloned specific, random or universal sequences that can detect restriction site heterogeneity in the target DNA. The detection of variation in rDNA gene loci is the basis of ribotyping , and this method has been universally applied to the typing of various species. Other commonly used probes are insertion sequences (lengths of DNA involved in transposition; see Ch. 6) that may define clonal structures of populations.
Polymerase chain reaction typing
PCR is a technique that allows specific sequences of DNA to be amplified. Multiple copies of regions of the genome defined by specific oligonucleotide primers are made by repeated cycles of amplification under controlled conditions. Such methods can be used to study DNA from any source. Several variations on the PCR theme have been described, and use of these techniques continues to expand and develop. PCR-mediated DNA fingerprinting makes use of the variable regions in DNA molecules. These may be variable numbers of tandem repeat regions or areas with restriction endonuclease recognition sequences. To perform PCR typing, it is necessary to know the sequences of the bordering regions so that specific oligonucleotide primers can be synthesized. Primers may be specific for a known sequence or be random. Random primers are extensively used in the techniques of random amplification of polymorphic DNA (RAPD) and arbitrarily primed PCR (AP-PCR). Both of these approaches have problems with reproducibility as a result of false priming, faint versus sharp bands and variation in electrophoretic migration of products. Repetitive sequence-based PCR (rep-PCR) indexes variation in multiple interspersed repetitive sequences in intergenic regions dispersed throughout the genome. An automated, standardized rep-PCR system has proven useful for strain typing of a number of species and is reported to give similar discrimination to PFGE (Bacterial Barcodes, Houston, Texas, USA). Amplified fragment length polymorphism is a DNA sequence-based technique that combines restriction endonuclease digestion with PCR. Incorporation of a fluorescent label and the use of a capillary DNA sequencer allows optimal standardization of reproducibility and resolution of single base-pair differences between genomes.
Multilocus sequence typing
This technique indexes allelic variation in several housekeeping genes by nucleotide sequencing rather than indirectly from the electrophoretic mobilities of their gene products, as was the case with its parent technique, multilocus enzyme electrophoresis. Housekeeping genes are not subject to selective forces as are variable genes and they diversify slowly. Multiple genes (usually seven) are employed to overcome the effects of recombination in a single locus, which might distort the interpretation of the relationship of the strains being compared. Multilocus sequence typing (MLST) can rightly be referred to as definitive genotyping as sequence data are unambiguous and databases of allelic profiles of isolates of individual species are accessible via the internet. The level of discrimination of MLST depends on the degree of diversity within the population to generate alleles at each locus, but some highly uniform species such as M. tuberculosis are not amenable to analysis by the technique. Recently, increased discrimination has been sought in virulence-associated genes necessary for survival and spread of the organism on the basis that these genes are exposed to frequent environmental changes and thus provide a higher degree of sequence variation. Intergenic regions of selected genes are amplified by PCR and a 500-bp internal fragment sequenced to identify allelic polymorphisms.
A variant of MLST termed multilocus restriction typing introduces restriction digestion of amplified housekeeping genes and removes the need for sequencing. The restriction fragment length polymorphisms (RFLPs) can be sorted into type patterns and reveal population structures similar to those with MLST.
Variable number tandem repeat analysis
Variable number tandem repeats (VNTRs) are short nucleotide sequences (20–100 bp) that vary in copy number in bacterial genomes. They are thought to arise through DNA strand slippage during replication and are of unknown function. Separate VNTR loci are identified from published sequences and are often located in intergenic regions and annotated open reading frames. Primers are designed to amplify five to eight loci and the products sequenced to generate a digital profile. VNTR typing is rapid and reproducible, and relatively simple to perform. Improved discrimination may be achieved by identification of more loci but there is debate about their stability over time.
Whole genome based typing
The advent of new high throughput DNA sequencing methods is ushering in a new era in which it is becoming feasible to compare and type bacteria based on entire genome sequence data. These massively parallel sequencing technologies produce relatively short nucleotide sequence reads but on such a scale that these can be assembled into a sequence matched against those obtained from previous isolates of that organism. This enables a genome-wide comparison to be made and a more or less definitive evolutionary relationship to be established to other contemporaneous and historical isolates. The costs of such analyses are rapidly becoming competitive with traditional typing methods. Such analyses have the potential to transform medical bacteriology by producing unambiguous epidemiological information and by identifying genetic elements such as those encoding antibiotic resistance and significant antigens under selection pressures.
Recommended reading
- Barrow GI, Feltham RKA, editors. Cowan and Steel's Manual for the Identification of Medical Bacteria. ed 3. Cambridge University Press; Cambridge: 1993. [ Google Scholar ]
- Garrity GM, editor. Bergey's Manual of Systematic Bacteriology. ed 2. Springer; New York: 2005. [ Google Scholar ]
- Kaufmann ME. Pulsed-field gel electrophoresis. In: Woodford N, Johnson AP, editors. Methods in Molecular Medicine , Vol. 15: Molecular Bacteriology: Protocols and Clinical Applications . Humana Press; Totowa, NJ: 1998. pp. 33–50. [ Google Scholar ]
- Murray PR, Baron EJ, Jorgensen JH, editors. Manual of Clinical Microbiology. ed 8. ASM Press; Washington, DC: 2003. [ Google Scholar ]
- Owen RJ. Bacterial taxonomics: finding the wood through the phylogenetic trees. Methods in Molecular Biology. 2004; 266 :353–384. [ PubMed ] [ Google Scholar ]
- Schleifer KH. Classification of Bacteria and Archeaea : Past, present and future. Systematic and Applied Microbiology. 2009; 32 :533–542. [ PubMed ] [ Google Scholar ]
- Spratt BG, Feil EJ, Smith NH. Population genetics of bacterial pathogens. In: Sussman M, editor. Molecular Medical Microbiology. Academic Press; San Diego: 2002. pp. 445–484. [ Google Scholar ]
- Tenover FC, Arbeit RD, Goering RV. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for strain typing. Journal of Clinical Microbiology. 1995; 33 :2233–2239. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Van Regenmortel MHV, Fauquet CM, Bishop DHL, editors. Virus Taxonomy. Classification and Nomenclature of Viruses. Academic Press; San Diego: 2000. [ Google Scholar ]
- Woese CR. Interpreting the universal phylogenetic tree. Proceedings of the National Academy of Sciences of the USA. 2000; 97 :8392–8396. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- http://www.mlst.net/ Genotyping database at Oxford University.
- http://www.ncbi.nlm.nih.gov/Genbank/ National Center for Biotechnology information for rRNA sequence analysis.
- http://www.ridom-rdna.de/ Ribosomal differentiation of medical microorganisms for rRNA sequence analysis.
- http://www.ictvdb.org/ Universal virus database of the International Committee on Taxonomy of Viruses.
IMAGES
VIDEO
COMMENTS
Nutrition in Bacteria: Definition. Nutrition is a process of acquiring energy and food. The need for energy is required for growth, reproduction, and maintenance. The main sources of nutrients in bacteria are carbon, nitrogen, water, phosphorous, iron, and some inorganic salts. Following are types of Bacteria.
Nutrition in Bacteria. Nutrition in bacteria is primarily categorized into two major groups, namely autotrophic and heterotrophic. Classification of bacteria in terms of nutritional requirement typically depends on the energy sources, whether in the form of light or chemical energy. Some bacteria obtain energy directly from the sunlight.
Heterotrophic Bacteria. The heterotrophic bacteria obtain their-ready made food from organic substances, living or dead. Most of pathogenic bacteria of human beings, other plants and animals are heterotrophs.; Some heterotrops have simple nutritional requirement while some of them require large amount of vitamin and other growth promoting substance. Such organisms are called fastidious ...
In conclusion, bacterial nutrition is a complex and diverse field that encompasses a wide array of metabolic strategies and ecological interactions. Autotrophic bacteria harness energy from inorganic sources, while heterotrophic bacteria rely on organic compounds. Mixotrophic bacteria combine both autotrophic and heterotrophic modes of nutrition
This page titled 11: Microbial Nutrition is shared under a CC BY-NC-SA license and was authored, remixed, and/or curated by Linda Bruslind ( Open Oregon State) . All microbes have a need for three things: carbon, energy, and electrons. There are specific terms associated with the source of each of these items, to help define organisms.
3.1 INTRODUCTION. Bacteria are prokaryotic organisms that do not contain chlorophyll. They are unicellular and do not show true branching. They differ from eukaryotes in not having a nuclear membrane, a nucleolus, and cell organelles like mitochondria, golgi apparatus and endoplasmic reticulum. They have a single circular chromosome.
Iron is required by microbes for the function of their cytochromes and enzymes, resulting in it being a growth-limiting micronutrient. However, little free iron is available in environments, due to its insolubility. Many bacteria have evolved siderophores, organic molecules that chelate or bind ferric iron with high affinity.
This nutritional mode is very common among eukaryotes, including humans. Photoautotrophs are cells that capture light energy, and use carbon dioxide as their carbon source. There are many photoautotrophic prokaryotes, which include cyanobacteria. ... This page titled 7.12: Bacteria Nutrition is shared under a CK-12 license and was authored, ...
Covers the different ways bacteria obtain energy and nutrients. Click Create Assignment to assign this modality to your LMS. We have a new and improved read on this topic.
Nutritional Classification of Bacteria. Table of Contents. Bacteria are prokaryotic organisms that require nutrients and energy for their growth and development like other living organisms. They require carbon, hydrogen, oxygen, nitrogen, metals and water for their biochemical processes. On the basis of their energy source and the carbon ...
Explore the mode of nutrition in bacteria. Discover how bacteria obtain energy and understand the classification of bacteria based on their energy sources. Updated: 10/13/2022
The autotrophic bacteria resemble green plants in their mode of nutrition but with a difference. The green plants utilize water as one of the raw materials in photosynthesis (food manufacturing process). The autotrophic bacteria, in contrast use, other compounds of hydrogen such as hydrogen sulphide (H 2 S) and methane (CH 4) and not water with ...
Biofilms are common among many types of bacteria. They rely on quorum sensing, a mechanism that bacteria use to "count" themselves to initiate a genetic response to their environment. The counting molecule is called an autoinducer. Bacteria use this molecule to help establish biofilms (fig. 7.14). 7.6 The Study of Microbial Growth
These are bacteria that live in the intestines of animals. They are usually beneficial bacteria, needed by the host organism. These microbes obviously don't kill their host, as that would kill the bacteria as well. Figure 5.3.1 5.3. 1: These mutualistic bacteria-containing nodules on a soybean root help provide the plant with nitrogen.
Carbon is often provided in carbohydrate form (glucose, lactose, mannitol, etc.) or as protein or protein breakdown products (peptones, peptides or proteoses). Media without carbon can be used to grow cyanobacteria. Nitrogen - Nitrogen is essential for the synthesis of proteins, nucleic acids and some carbohydrates, e.g., glucosamine.
Autotrophic Nutrition. In the autotrophic mode, organisms use simple inorganic matters like water and carbon dioxide in the presence of light and chlorophyll to synthesize food on their own. In other words, the process of photosynthesis is used to convert light energy into food such as glucose. Such organisms are called autotrophs.
The LibreTexts libraries are Powered by NICE CXone Expert and are supported by the Department of Education Open Textbook Pilot Project, the UC Davis Office of the Provost, the UC Davis Library, the California State University Affordable Learning Solutions Program, and Merlot. We also acknowledge previous National Science Foundation support under grant numbers 1246120, 1525057, and 1413739.
Energy can be tapped from light or chemicals (organic chemicals and inorganic chemicals). The nutritional types of bacteria vary greatly. Organisms that can utilize radiant energy (sunlight) are called phototrophs. Chemotrophs are organisms that can harvest energy from chemicals. Chemotrophs. Chemoorganotrophs. Chemolithotrophs. Photolithotrophs.
The microbial cell is made up of several elements such as carbon, oxygen, hydrogen, nitrogen, sulfur, phosphorus, potassium, calcium, magnesium, and iron. These are also known as macro elements or macronutrients because these elements are required in high amounts by the microbes. Among these, C, H, O, N, S, and P are the major elements required ...
Bacteria can be classified based on shape, mode of nutrition, respiration, the composition of the cell wall, etc. Based on these criteria, bacteria can be classified as bacillus, coccus or vibrio, etc., autotrophic or heterotrophic, aerobic or anaerobic, Gram + or Gram -. Read the above topic for detailed information.
This page titled 4.2: Microbial Nutrition is shared under a CC BY-NC-SA license and was authored, remixed, and/or curated by Linda Bruslind ( Open Oregon State) . All microbes have a need for three things: carbon, energy, and electrons. There are specific terms associated with the source of each of these items, to help define organisms.
environment. Some bacteria have a generation time of 24 hours, although the generation time of most bacteria is between 1 to 3 hours. • Bacterial cells grow at an enormous rate. For example, with binary fission, bacteria can double every 20 minutes. In 30 generations of bacteria (10 hours), the number could reach one billion. It is difficult
The algae (excluding the blue-green algae), the protozoa, slime moulds and fungi include the larger eukaryotic (see Ch. 2) micro-organisms; their cells have the same general type of structure and organization as that found in plants and animals. The bacteria, including organisms of the mycoplasma, rickettsia and chlamydia groups, together with the related blue-green algae, comprise the ...
We go hands-on with the Sonos Ace, the company's long awaited headphones debut that offers a stunning design, impressive sound and compatibility with Sonos soundbars.
Instead of announcing the Surface Pro 10 or Surface Laptop 6, Microsoft revealed reborn Copilot+ PC versions with seriously upgraded performance and battery life.