Thank you for visiting nature.com. You are using a browser version with limited support for CSS. To obtain the best experience, we recommend you use a more up to date browser (or turn off compatibility mode in Internet Explorer). In the meantime, to ensure continued support, we are displaying the site without styles and JavaScript.
- View all journals
- Explore content
- About the journal
- Publish with us
- Sign up for alerts
- Review Article
- Published: 27 June 2019

The human stress response
- Georgina Russell 1 &
- Stafford Lightman ORCID: orcid.org/0000-0002-8546-9646 1
Nature Reviews Endocrinology volume 15 , pages 525–534 ( 2019 ) Cite this article
25k Accesses
431 Citations
833 Altmetric
Metrics details
- Adrenal cortex hormones
- Circadian rhythms
- Multihormonal system disorders
- Stress signalling
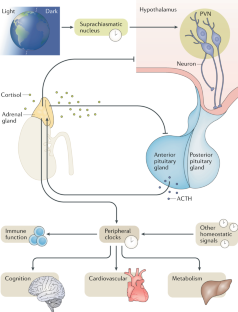
The human stress response has evolved to maintain homeostasis under conditions of real or perceived stress. This objective is achieved through autoregulatory neural and hormonal systems in close association with central and peripheral clocks. The hypothalamic–pituitary–adrenal axis is a key regulatory pathway in the maintenance of these homeostatic processes. The end product of this pathway — cortisol — is secreted in a pulsatile pattern, with changes in pulse amplitude creating a circadian pattern. During acute stress, cortisol levels rise and pulsatility is maintained. Although the initial rise in cortisol follows a large surge in adrenocorticotropic hormone levels, if long-term inflammatory stress occurs, adrenocorticotropic hormone levels return to near basal levels while cortisol levels remain raised as a result of increased adrenal sensitivity. In chronic stress, hypothalamic activation of the pituitary changes from corticotropin-releasing hormone-dominant to arginine vasopressin-dominant, and cortisol levels remain raised due at least in part to decreased cortisol metabolism. Acute elevations in cortisol levels are beneficial to promoting survival of the fittest as part of the fight-or-flight response. However, chronic exposure to stress results in reversal of the beneficial effects, with long-term cortisol exposure becoming maladaptive, which can lead to a broad range of problems including the metabolic syndrome, obesity, cancer, mental health disorders, cardiovascular disease and increased susceptibility to infections. Neuroimmunoendocrine modulation in disease states and glucocorticoid-based therapeutics are also discussed.
The hypothalamic–pituitary–adrenal (HPA) axis is a key system that synchronizes the stress response with circadian regulatory processes.
Regulation of the HPA axis is very dynamic with both ultradian and circadian oscillations.
Short-term and longer-term stress result in different regulatory mechanisms involving hypothalamic, pituitary and adrenal activity, as well as cortisol metabolism.
Chronic elevation and nonphysiological patterns of cortisol result in poor cognitive, metabolic and immune function.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
24,99 € / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
195,33 € per year
only 16,28 € per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout
Similar content being viewed by others

The neuroendocrinology of stress: the stress-related continuum of chronic disease development

Systematic manipulations of the biological stress systems result in sex-specific compensatory stress responses and negative mood outcomes

The cortisol switch between vulnerability and resilience
Szabo, S., Tache, Y. & Somogyi, A. The legacy of Hans Selye and the origins of stress research: a retrospective 75 years after his landmark brief “letter” to the editor of Nature . Stress 15 , 472–478 (2012).
CAS PubMed Google Scholar
Levine, S. Influence of psychological variables on the activity of the hypothalamic-pituitary-adrenal axis. Eur. J. Pharmacol. 405 , 149–160 (2000).
Brown, S. A. & Azzi, A. Peripheral circadian oscillators in mammals. Handb. Exp. Pharmacol. 2013 , 45–66 (2013).
Google Scholar
Roenneberg, T. & Merrow, M. The circadian clock and human health. Curr. Biol. 26 , R432–R443 (2016).
Bass, J. & Lazar, M. A. Circadian time signatures of fitness and disease. Science 354 , 994–999 (2016).
Turek, F. W. Circadian neural rhythms in mammals. Annu. Rev. Physiol. 47 , 49–64 (1985).
Skene, D. J. et al. Separation of circadian- and behavior-driven metabolite rhythms in humans provides a window on peripheral oscillators and metabolism. Proc. Natl Acad. Sci. USA 115 , 7825–7830 (2018).
Buhr, E. D. & Takahashi, J. S. Molecular components of the mammalian circadian clock. Handb. Exp. Pharmacol. 217 , 3–27 (2013).
CAS Google Scholar
Takahashi, J. S. Transcriptional architecture of the mammalian circadian clock. Nat. Rev. Genet. 18 , 164–179 (2017).
Baron, K. G. & Reid, K. J. Circadian misalignment and health. Int. Rev. Psychiatry 26 , 139–154 (2014).
PubMed PubMed Central Google Scholar
Potter, G. D. et al. Circadian rhythm and sleep disruption: causes, metabolic consequences, and countermeasures. Endocr. Rev. 37 , 584–608 (2016).
CAS PubMed PubMed Central Google Scholar
Zhang, R., Lahens, N. F., Ballance, H. I., Hughes, M. E. & Hogenesch, J. B. A circadian gene expression atlas in mammals: implications for biology and medicine. Proc. Natl Acad. Sci. USA 111 , 16219–16224 (2014).
Smarr, B. L. & Schirmer, A. E. 3.4 million real-world learning management system logins reveal the majority of students experience social jet lag correlated with decreased performance. Sci. Rep. 8 , 4793 (2018).
Gardner, M. et al. Dysregulation of the hypothalamic pituitary adrenal (HPA) axis and cognitive capability at older ages: individual participant meta-analysis of five cohorts. Sci. Rep. 9 , 4555 (2019).
Selye, H. Stress and the general adaptation syndrome. BMJ 1 , 1383–1392 (1950).
Russell, G. M. & Lightman, S. L. Can side effects of steroid treatments be minimized by the temporal aspects of delivery method? Expert Opin. Drug Saf. 13 , 1501–1513 (2014).
Sorrells, S. F. & Sapolsky, R. M. An inflammatory review of glucocorticoid actions in the CNS. Brain Behav. Immun. 21 , 259–272 (2007).
Busillo, J. M. & Cidlowski, J. A. The five Rs of glucocorticoid action during inflammation: ready, reinforce, repress, resolve, and restore. Trends Endocrinol. Metab. 24 , 109–119 (2013).
McEwen, B. S. et al. The role of adrenocorticoids as modulators of immune function in health and disease: neural, endocrine and immune interactions. Brain Res. Rev. 23 , 79–133 (1997).
Elenkov, I. J. & Chrousos, G. P. Stress system — organization, physiology and immunoregulation. Neuroimmunomodulation 13 , 257–267 (2006).
Brinkmann, V. & Kristofic, C. Regulation by corticosteroids of Th1 and Th2 cytokine production in human CD4+ effector T cells generated from CD45RO- and CD45RO+ subsets. J. Immunol. 155 , 3322–3328 (1995).
Wiegers, G. J. & Reul, J. M. Induction of cytokine receptors by glucocorticoids: functional and pathological significance. Trends Pharmacol. Sci. 19 , 317–321 (1998).
Abraham, I. M., Meerlo, P. & Luiten, P. G. Concentration dependent actions of glucocorticoids on neuronal viability and survival. Dose Response 4 , 38–54 (2006).
Plaschke, K., Muller, D. & Hoyer, S. Effect of adrenalectomy and corticosterone substitution on glucose and glycogen metabolism in rat brain. J. Neural Transm. 103 , 89–100 (1996).
Belanoff, J. K., Gross, K., Yager, A. & Schatzberg, A. F. Corticosteroids and cognition. J. Psychiatr. Res. 35 , 127–145 (2001).
Roozendaal, B. Stress and memory: opposing effects of glucocorticoids on memory consolidation and memory retrieval. Neurobiol. Learn. Mem. 78 , 578–595 (2002).
Brown, E. S. Effects of glucocorticoids on mood, memory, and the hippocampus. Treatment and preventive therapy. Ann. NY Acad. Sci. 1179 , 41–55 (2009).
de Kloet, E. R., Oitzl, M. S. & Joels, M. Stress and cognition: are corticosteroids good or bad guys? Trends Neurosci. 22 , 422–426 (1999).
PubMed Google Scholar
Decani, S., Federighi, V., Baruzzi, E., Sardella, A. & Lodi, G. Iatrogenic Cushing’s syndrome and topical steroid therapy: case series and review of the literature. J. Dermatolog. Treat. 25 , 495–500 (2014).
Kenna, H. A., Poon, A. W., de los Angeles, C. P. & Koran, L. M. Psychiatric complications of treatment with corticosteroids: review with case report. Psychiatry Clin. Neurosci. 65 , 549–560 (2011).
Buijs, R. M., Markman, M., Nunes-Cardoso, B., Hou, Y. X. & Shinn, S. Projections of the suprachiasmatic nucleus to stress-related areas in the rat hypothalamus: a light and electron microscopic study. J. Comp. Neurol. 335 , 42–54 (1993).
Watts, A. G. & Swanson, L. W. Efferent projections of the suprachiasmatic nucleus: II. Studies using retrograde transport of fluorescent dyes and simultaneous peptide immunohistochemistry in the rat. J. Comp. Neurol. 258 , 230–252 (1987).
Jacobson, L. Hypothalamic-pituitary-adrenocortical axis regulation. Endocrinol. Metab. Clin. North Am. 34 , 271–292 (2005).
Herman, J. P., Ostrander, M. M., Mueller, N. K. & Figueiredo, H. Limbic system mechanisms of stress regulation: hypothalamo-pituitary-adrenocortical axis. Prog. Neuropsychopharmacol. Biol. Psychiatry 29 , 1201–1213 (2005).
Dallman, M. F. et al. Corticosteroids and the control of function in the hypothalamo-pituitary-adrenal (HPA) axis. Ann. NY Acad. Sci. 746 , 22–31 (1994).
Jasper, M. S. & Engeland, W. C. Splanchnic neural activity modulates ultradian and circadian rhythms in adrenocortical secretion in awake rats. Neuroendocrinology 59 , 97–109 (1994).
Buijs, R. M. et al. Anatomical and functional demonstration of a multisynaptic suprachiasmatic nucleus adrenal (cortex) pathway. Eur. J. Neurosci. 11 , 1535–1544 (1999).
Kiessling, S., Sollars, P. J. & Pickard, G. E. Light stimulates the mouse adrenal through a retinohypothalamic pathway independent of an effect on the clock in the suprachiasmatic nucleus. PLOS ONE 9 , e92959 (2014).
Husse, J., Leliavski, A., Tsang, A. H., Oster, H. & Eichele, G. The light-dark cycle controls peripheral rhythmicity in mice with a genetically ablated suprachiasmatic nucleus clock. FASEB J. 28 , 4950–4960 (2014).
Ishida, A. et al. Light activates the adrenal gland: timing of gene expression and glucocorticoid release. Cell Metab. 2 , 297–307 (2005).
Oster, H. et al. The circadian rhythm of glucocorticoids is regulated by a gating mechanism residing in the adrenal cortical clock. Cell Metab. 4 , 163–173 (2006).
Charmandari, E. et al. Peripheral CLOCK regulates target-tissue glucocorticoid receptor transcriptional activity in a circadian fashion in man. PLOS ONE 6 , e25612 (2011).
Bailey, S. L. & Heitkemper, M. M. Circadian rhythmicity of cortisol and body temperature: morningness-eveningness effects. Chronobiol. Int. 18 , 249–261 (2001).
Donner, N. C., Montoya, C. D., Lukkes, J. L. & Lowry, C. A. Chronic non-invasive corticosterone administration abolishes the diurnal pattern of tph2 expression. Psychoneuroendocrinology 37 , 645–661 (2012).
Lightman, S. L. The neuroendocrinology of stress: a never ending story. J. Neuroendocrinol. 20 , 880–884 (2008).
Nicolaides, N. C., Kyratzi, E., Lamprokostopoulou, A., Chrousos, G. P. & Charmandari, E. Stress, the stress system and the role of glucocorticoids. Neuroimmunomodulation 22 , 6–19 (2015).
Gibbison, B. et al. Dynamic pituitary-adrenal interactions in response to cardiac surgery. Crit. Care Med. 43 , 791–800 (2015).
Spiga, F. et al. Dynamic responses of the adrenal steroidogenic regulatory network. Proc. Natl Acad. Sci. USA 114 , E6466–E6474 (2017).
Ma, X. M., Levy, A. & Lightman, S. L. Emergence of an isolated arginine vasopressin (AVP) response to stress after repeated restraint: a study of both AVP and corticotropin-releasing hormone messenger ribonucleic acid (RNA) and heteronuclear RNA. Endocrinology 138 , 4351–4357 (1997).
Dallman, M. F. Stress update: adaptation of the hypothalamic-pituitary-adrenal axis to chronic stress. Trends. Endocrinol. Metab. 4 , 62–69 (1993).
Henley, D. E. et al. Hypothalamic-pituitary-adrenal axis activation in obstructive sleep apnea: the effect of continuous positive airway pressure therapy. J. Clin. Endocrinol. Metab. 94 , 4234–4242 (2009).
Boonen, E. et al. Reduced cortisol metabolism during critical illness. N. Engl. J. Med. 368 , 1477–1488 (2013).
Peeters, B. et al. Adrenocortical function during prolonged critical illness and beyond: a prospective observational study. Intensive Care Med. 44 , 1720–1729 (2018).
De Kloet, E. R., Vreugdenhil, E., Oitzl, M. S. & Joels, M. Brain corticosteroid receptor balance in health and disease. Endocr. Rev. 19 , 269–301 (1998).
Reul, J. M. & de Kloet, E. R. Two receptor systems for corticosterone in rat brain: microdistribution and differential occupation. Endocrinology 117 , 2505–2511 (1985).
Dallman, M. F. Fast glucocorticoid actions on brain: back to the future. Front. Neuroendocrinol. 26 , 103–108 (2005).
Russell, G. M. et al. Rapid glucocorticoid receptor-mediated inhibition of hypothalamic-pituitary-adrenal ultradian activity in healthy males. J. Neurosci. 30 , 6106–6115 (2010).
Lowenberg, M., Verhaar, A. P., van den Brink, G. R. & Hommes, D. W. Glucocorticoid signaling: a nongenomic mechanism for T cell immunosuppression. Trends Mol. Med. 13 , 158–163 (2007).
Orchinik, M., Murray, T. F., Franklin, P. H. & Moore, F. L. Guanyl nucleotides modulate binding to steroid receptors in neuronal membranes. Proc. Natl Acad. Sci. USA 89 , 3830–3834 (1992).
Orchinik, M., Murray, T. F. & Moore, F. L. A corticosteroid receptor in neuronal membranes. Science 252 , 1848–1851 (1991).
Joels, M., Pasricha, N. & Karst, H. The interplay between rapid and slow corticosteroid actions in brain. Eur. J. Pharmacol. 719 , 44–52 (2013).
Walker, J. J. et al. The origin of glucocorticoid hormone oscillations. PLOS Biol. 10 , e1001341 (2012).
Patel, P. D. et al. Glucocorticoid and mineralocorticoid receptor mRNA expression in squirrel monkey brain. J. Psychiatr. Res. 34 , 383–392 (2000).
Groeneweg, F. L., Karst, H., de Kloet, E. R. & Joels, M. Rapid non-genomic effects of corticosteroids and their role in the central stress response. J. Endocrinol. 209 , 153–167 (2011).
de Kloet, E. R., Fitzsimons, C. P., Datson, N. A., Meijer, O. C. & Vreugdenhil, E. Glucocorticoid signaling and stress-related limbic susceptibility pathway: about receptors, transcription machinery and microRNA. Brain Res. 1293 , 129–141 (2009).
Russell, G. M., Kalafatakis, K. & Lightman, S. L. The importance of biological oscillators for HPA activity and tissue glucocorticoid response: coordinating stress and neurobehavioural adaptation. J. Neuroendocrinol. 27 , 378–388 (2015).
Lewis, J. G. et al. Plasma variation of corticosteroid-binding globulin and sex hormone-binding globulin. Horm. Metab. Res. 38 , 241–245 (2006).
Lewis, J. G., Bagley, C. J., Elder, P. A., Bachmann, A. W. & Torpy, D. J. Plasma free cortisol fraction reflects levels of functioning corticosteroid-binding globulin. Clin. Chim. Acta 359 , 189–194 (2005).
Hammond, G. L., Smith, C. L. & Underhill, D. A. Molecular studies of corticosteroid binding globulin structure, biosynthesis and function. J. Steroid Biochem. Mol. Biol. 40 , 755–762 (1991).
Frairia, R. et al. Influence of naturally occurring and synthetic glucocorticoids on corticosteroid-binding globulin-steroid interaction in human peripheral plasma. Ann. NY Acad. Sci. 538 , 287–303 (1988).
Cameron, A. et al. Temperature-responsive release of cortisol from its binding globulin: a protein thermocouple. J. Clin. Endocrinol. Metab. 95 , 4689–4695 (2010).
Kyrou, I., Chrousos, G. P. & Tsigos, C. Stress, visceral obesity, and metabolic complications. Ann. NY Acad. Sci. 1083 , 77–110 (2006).
Chapman, K., Holmes, M. & Seckl, J. 11beta-hydroxysteroid dehydrogenases: intracellular gate-keepers of tissue glucocorticoid action. Physiol. Rev. 93 , 1139–1206 (2013).
Seckl, J. R. 11beta-hydroxysteroid dehydrogenases: changing glucocorticoid action. Curr. Opin. Pharmacol. 4 , 597–602 (2004).
Verma, M. et al. 11beta-hydroxysteroid dehydrogenase-1 deficiency alters brain energy metabolism in acute systemic inflammation. Brain Behav. Immun. 69 , 223–234 (2018).
Follenius, M., Simon, C., Brandenberger, G. & Lenzi, P. Ultradian plasma corticotropin and cortisol rhythms: time-series analyses. J. Endocrinol. Invest. 10 , 261–266 (1987).
Hartmann, A., Veldhuis, J. D., Deuschle, M., Standhardt, H. & Heuser, I. Twenty-four hour cortisol release profiles in patients with Alzheimer’s and Parkinson’s disease compared to normal controls: ultradian secretory pulsatility and diurnal variation. Neurobiol. Aging 18 , 285–289 (1997).
Rivest, R. W., Schulz, P., Lustenberger, S. & Sizonenko, P. C. Differences between circadian and ultradian organization of cortisol and melatonin rhythms during activity and rest. J. Clin. Endocrinol. Metab. 68 , 721–729 (1989).
Waite, E. J. et al. Ultradian corticosterone secretion is maintained in the absence of circadian cues. Eur. J. Neurosci. 36 , 3142–3150 (2012).
Ixart, G., Barbanel, G., Nouguier-Soule, J. & Assenmacher, I. A quantitative study of the pulsatile parameters of CRH-41 secretion in unanesthetized free-moving rats. Exp. Brain Res. 87 , 153–158 (1991).
Spiga, F. et al. ACTH-dependent ultradian rhythm of corticosterone secretion. Endocrinology 152 , 1448–1457 (2011).
Spiga, F., Liu, Y., Aguilera, G. & Lightman, S. L. Temporal effect of adrenocorticotrophic hormone on adrenal glucocorticoid steroidogenesis: involvement of the transducer of regulated cyclic AMP-response element-binding protein activity. J. Neuroendocrinol. 23 , 136–142 (2011).
Lim, C. & Allada, R. Emerging roles for post-transcriptional regulation in circadian clocks. Nat. Neurosci. 16 , 1544–1550 (2013).
Liston, C. et al. Circadian glucocorticoid oscillations promote learning-dependent synapse formation and maintenance. Nat. Neurosci. 16 , 698–705 (2013).
Lightman, S. L. & Conway-Campbell, B. L. The crucial role of pulsatile activity of the HPA axis for continuous dynamic equilibration. Nat. Rev. Neurosci. 11 , 710–718 (2010).
Stavreva, D. A. et al. Ultradian hormone stimulation induces glucocorticoid receptor-mediated pulses of gene transcription. Nat. Cell Biol. 11 , 1093–1102 (2009).
Conway-Campbell, B. L., Pooley, J. R., Hager, G. L. & Lightman, S. L. Molecular dynamics of ultradian glucocorticoid receptor action. Mol. Cell. Endocrinol. 348 , 383–393 (2012).
George, C. L., Lightman, S. L. & Biddie, S. C. Transcription factor interactions in genomic nuclear receptor function. Epigenomics 3 , 471–485 (2011).
So, A. Y., Chaivorapol, C., Bolton, E. C., Li, H. & Yamamoto, K. R. Determinants of cell- and gene-specific transcriptional regulation by the glucocorticoid receptor. PLOS Genet. 3 , e94 (2007).
Zalachoras, I., Houtman, R. & Meijer, O. C. Understanding stress-effects in the brain via transcriptional signal transduction pathways. Neuroscience 242 , 97–109 (2013).
Sarabdjitsingh, R. A. et al. Stress responsiveness varies over the ultradian glucocorticoid cycle in a brain-region-specific manner. Endocrinology 151 , 5369–5379 (2010).
Sarabdjitsingh, R. A. et al. Ultradian corticosterone pulses balance glutaminergic transmission and synaptic plasticity. Proc. Natl Acad. Sci. USA 111 , 14265–14270 (2014).
Kalafatakis, K. et al. Ultradian rhythmicity of plasma cortisol is necessary for normal emotional and cognitive responses in man. Proc. Natl Acad. Sci. USA 115 , E4091–E4100 (2018).
Gjerstad, J. K., Lightman, S. L. & Spiga, F. Role of glucocorticoid negative feedback in the regulation of HPA axis pulsatility. Stress 21 , 403–416 (2018).
Bornstein, S. R., Engeland, W. C., Ehrhart-Bornstein, M. & Herman, J. P. Dissociation of ACTH and glucocorticoids. Trends Endocrinol. Metab. 19 , 175–180 (2008).
Silverman, M. N., Miller, A. H., Biron, C. A. & Pearce, B. D. Characterization of an interleukin-6- and adrenocorticotropin-dependent, immune-to-adrenal pathway during viral infection. Endocrinology 145 , 3580–3589 (2004).
Franchimont, D. et al. Adrenal cortical activation in murine colitis. Gastroenterology 119 , 1560–1568 (2000).
Viblanc, V. A. et al. An integrative appraisal of the hormonal and metabolic changes induced by acute stress using king penguins as a model. Gen. Comp. Endocrinol. 269 , 1–10 (2018).
Cruz-Topete, D. & Cidlowski, J. A. One hormone, two actions: anti- and pro-inflammatory effects of glucocorticoids. Neuroimmunomodulation 22 , 20–32 (2015).
Biddie, S. C., Conway-Campbell, B. L. & Lightman, S. L. Dynamic regulation of glucocorticoid signalling in health and disease. Rheumatology 51 , 403–412 (2012).
Miller, G. E., Cohen, S. & Ritchey, A. K. Chronic psychological stress and the regulation of pro-inflammatory cytokines: a glucocorticoid-resistance model. Health Psychol. 21 , 531–541 (2002).
Oster, H. et al. The functional and clinical significance of the 24-hour rhythm of circulating glucocorticoids. Endocr. Rev. 38 , 3–45 (2017).
Keller, M. et al. A circadian clock in macrophages controls inflammatory immune responses. Proc. Natl Acad. Sci. USA 106 , 21407–21412 (2009).
Boivin, D. B. et al. Circadian clock genes oscillate in human peripheral blood mononuclear cells. Blood 102 , 4143–4145 (2003).
Koo, J. W., Russo, S. J., Ferguson, D., Nestler, E. J. & Duman, R. S. Nuclear factor-kappaB is a critical mediator of stress-impaired neurogenesis and depressive behavior. Proc. Natl Acad. Sci. USA 107 , 2669–2674 (2010).
Pace, T. W., Hu, F. & Miller, A. H. Cytokine-effects on glucocorticoid receptor function: relevance to glucocorticoid resistance and the pathophysiology and treatment of major depression. Brain Behav. Immun. 21 , 9–19 (2007).
Pace, T. W. et al. Increased stress-induced inflammatory responses in male patients with major depression and increased early life stress. Am. J. Psychiatry 163 , 1630–1633 (2006).
Cohen, S. et al. Chronic stress, glucocorticoid receptor resistance, inflammation, and disease risk. Proc. Natl Acad. Sci. USA 109 , 5995–5999 (2012).
Spiegel, K., Leproult, R. & Van Cauter, E. Impact of sleep debt on metabolic and endocrine function. Lancet 354 , 1435–1439 (1999).
Hauner, H., Schmid, P. & Pfeiffer, E. F. Glucocorticoids and insulin promote the differentiation of human adipocyte precursor cells into fat cells. J. Clin. Endocrinol. Metab. 64 , 832–835 (1987).
Dallman, M. F. et al. Glucocorticoids, chronic stress, and obesity. Prog. Brain Res. 153 , 75–105 (2006).
Tsigos, C. et al. Dose-dependent effects of recombinant human interleukin-6 on glucose regulation. J. Clin. Endocrinol. Metab. 82 , 4167–4170 (1997).
McEwen, B. S. Sleep deprivation as a neurobiologic and physiologic stressor: allostasis and allostatic load. Metabolism 55 , S20–S23 (2006).
Zhu, B., Shi, C., Park, C. G., Zhao, X. & Reutrakul, S. Effects of sleep restriction on metabolism-related parameters in healthy adults: a comprehensive review and meta-analysis of randomized controlled trials. Sleep Med. Rev. 45 , 18–30 (2019).
Gavrila, A. et al. Diurnal and ultradian dynamics of serum adiponectin in healthy men: comparison with leptin, circulating soluble leptin receptor, and cortisol patterns. J. Clin. Endocrinol. Metab. 88 , 2838–2843 (2003).
Knutson, K. L. & Van Cauter, E. Associations between sleep loss and increased risk of obesity and diabetes. Ann. NY Acad. Sci. 1129 , 287–304 (2008).
Adam, T. C. & Epel, E. S. Stress, eating and the reward system. Physiol. Behav. 91 , 449–458 (2007).
Young, E. A., Carlson, N. E. & Brown, M. B. Twenty-four-hour ACTH and cortisol pulsatility in depressed women. Neuropsychopharmacology 25 , 267–276 (2001).
Heuser, I., Yassouridis, A. & Holsboer, F. The combined dexamethasone/CRH test: a refined laboratory test for psychiatric disorders. J. Psychiatr. Res. 28 , 341–356 (1994).
Ising, M. et al. The combined dexamethasone/CRH test as a potential surrogate marker in depression. Prog. Neuropsychopharmacol. Biol. Psychiatry 29 , 1085–1093 (2005).
Krishnan, V. & Nestler, E. J. The molecular neurobiology of depression. Nature 455 , 894–902 (2008).
Miller, A. H. Depression and immunity: a role for T cells? Brain Behav. Immun. 24 , 1–8 (2010).
Koo, J. W. & Duman, R. S. IL-1beta is an essential mediator of the antineurogenic and anhedonic effects of stress. Proc. Natl Acad. Sci. USA 105 , 751–756 (2008).
Horowitz, M. A., Zunszain, P. A., Anacker, C., Musaelyan, K. & Pariante, C. M. Glucocorticoids and inflammation: a double-headed sword in depression? How do neuroendocrine and inflammatory pathways interact during stress to contribute to the pathogenesis of depression? Mod. Trends Pharmacopsychiatry 28 , 127–143 (2013).
Munhoz, C. D. et al. Chronic unpredictable stress exacerbates lipopolysaccharide-induced activation of nuclear factor-kappaB in the frontal cortex and hippocampus via glucocorticoid secretion. J. Neurosci. 26 , 3813–3820 (2006).
Pariante, C. M. Glucocorticoid receptor function in vitro in patients with major depression. Stress 7 , 209–219 (2004).
Kenis, G. & Maes, M. Effects of antidepressants on the production of cytokines. Int. J. Neuropsychopharmacol. 5 , 401–412 (2002).
Bjornsdottir, S. et al. Drug prescription patterns in patients with Addison’s disease: a Swedish population-based cohort study. J. Clin. Endocrinol. Metab. 98 , 2009–2018 (2013).
Dunlop, D. Eighty-six cases of Addison’s disease. BMJ 2 , 887–891 (1963).
Giordano, R. et al. Metabolic and cardiovascular profile in patients with Addison’s disease under conventional glucocorticoid replacement. J. Endocrinol. Invest. 32 , 917–923 (2009).
Johannsson, G. et al. Adrenal insufficiency: review of clinical outcomes with current glucocorticoid replacement therapy. Clin. Endocrinol. 82 , 2–11 (2015).
Lovas, K., Loge, J. H. & Husebye, E. S. Subjective health status in Norwegian patients with Addison’s disease. Clin. Endocrinol. 56 , 581–588 (2002).
Feek, C. M. et al. Patterns of plasma cortisol and ACTH concentrations in patients with Addison’s disease treated with conventional corticosteroid replacement. Clin. Endocrinol. 14 , 451–458 (1981).
Isidori, A. M. et al. Effect of once-daily, modified-release hydrocortisone versus standard glucocorticoid therapy on metabolism and innate immunity in patients with adrenal insufficiency (DREAM): a single-blind, randomised controlled trial. Lancet Diabetes Endocrinol. 6 , 173–185 (2018).
Bancos, I. et al. Primary adrenal insufficiency is associated with impaired natural killer cell function: a potential link to increased mortality. Eur. J. Endocrinol. 176 , 471–480 (2017).
Bjanesoy, T. E. et al. Altered DNA methylation profile in Norwegian patients with autoimmune Addison’s disease. Mol. Immunol. 59 , 208–216 (2014).
Langenheim, J., Ventz, M., Hinz, A. & Quinkler, M. Modified-release prednisone decreases complaints and fatigue compared to standard prednisolone in patients with adrenal insufficiency. Horm. Metab. Res. 45 , 96–101 (2013).
Mallappa, A. et al. A phase 2 study of Chronocort, a modified-release formulation of hydrocortisone, in the treatment of adults with classic congenital adrenal hyperplasia. J. Clin. Endocrinol. Metab. 100 , 1137–1145 (2015).
Lovas, K. & Husebye, E. S. Continuous subcutaneous hydrocortisone infusion in Addison’s disease. Eur. J. Endocrinol. 157 , 109–112 (2007).
Venneri, M. A. et al. Circadian rhythm of glucocorticoid administration entrains clock genes in immune cells: a DREAM trial ancillary study. J. Clin. Endocrinol. Metab. 103 , 2998–3009 (2018).
Oksnes, M. et al. Continuous subcutaneous hydrocortisone infusion versus oral hydrocortisone replacement for treatment of Addison’s disease: a randomized clinical trial. J. Clin. Endocrinol. Metab. 99 , 1665–1674 (2014).
Riedel, M., Wiese, A., Schurmeyer, T. H. & Brabant, G. Quality of life in patients with Addison’s disease: effects of different cortisol replacement modes. Exp. Clin. Endocrinol. 101 , 106–111 (1993).
van Staa, T. P. et al. Use of oral corticosteroids in the United Kingdom. QJM 93 , 105–111 (2000).
Overman, R. A., Yeh, J. Y. & Deal, C. L. Prevalence of oral glucocorticoid usage in the United States: a general population perspective. Arthritis Care Res. 65 , 294–298 (2013).
Curtis, J. R. et al. Population-based assessment of adverse events associated with long-term glucocorticoid use. Arthritis Rheum. 55 , 420–426 (2006).
McDonough, A. K., Curtis, J. R. & Saag, K. G. The epidemiology of glucocorticoid-associated adverse events. Curr. Opin. Rheumatol. 20 , 131–137 (2008).
Leung, D. Y. & Bloom, J. W. Update on glucocorticoid action and resistance. J. Allergy Clin. Immunol. 111 , 3–22 (2003).
Download references
Author information
Authors and affiliations.
Translational Health Sciences, Dorothy Hodgkin Building, Bristol Medical School, University of Bristol, Bristol, UK
Georgina Russell & Stafford Lightman
You can also search for this author in PubMed Google Scholar
Contributions
The authors contributed equally to all aspects of the article.
Corresponding authors
Correspondence to Georgina Russell or Stafford Lightman .
Ethics declarations
Competing interests.
The authors declare no competing interests.
Additional information
Publisher’s note.
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Cues that entrain or synchronize the body’s 24-h cycle
Biological rhythms that occur with a frequency of <24 h.
A biochemical oscillator with phases synchronized with solar time.
Neural pathways involving at least one relay.
The microcirculation that allows transport of hypothalamic hormones to the pituitary gland.
The threshold power of (solar) electromagnetic radiation needed to exert an effect.
Repetitive body movements that serve no biological function.
Behaviours engaged for a specific functional purpose.
Any biological process that displays an oscillation of approximately 24 h.
Rights and permissions
Reprints and permissions
About this article
Cite this article.
Russell, G., Lightman, S. The human stress response. Nat Rev Endocrinol 15 , 525–534 (2019). https://doi.org/10.1038/s41574-019-0228-0
Download citation
Accepted : 05 June 2019
Published : 27 June 2019
Issue Date : September 2019
DOI : https://doi.org/10.1038/s41574-019-0228-0
Share this article
Anyone you share the following link with will be able to read this content:
Sorry, a shareable link is not currently available for this article.
Provided by the Springer Nature SharedIt content-sharing initiative
This article is cited by
Key hpi axis receptors facilitate light adaptive behavior in larval zebrafish.
- Soaleha Shams
- Karl J. Clark
Scientific Reports (2024)
Evaluation of the benefits of neutral bicarbonate ionized water baths in an open-label, randomized, crossover trial
- Ryoko Ushikoshi-Nakayama
- Tomoe Yamazaki
- Ichiro Saito
Sex-specific associations of serum cortisol with brain biomarkers of Alzheimer’s risk
- Lisa Mosconi
- Schantel Williams
- Jonathan P. Dyke
Evaluation of indicators of acute emotional states in dogs
- Hannah E. Flint
- Jennifer E. Weller
- Tammie King
Early Childhood Education Teacher Workforce: Stress in Relation to Identity and Choices
- Cynthia A. Wiltshire
Early Childhood Education Journal (2024)
Quick links
- Explore articles by subject
- Guide to authors
- Editorial policies
Sign up for the Nature Briefing newsletter — what matters in science, free to your inbox daily.

Stress Research: Past, Present, and Future
- Reference work entry
- First Online: 18 October 2022
- pp 2717–2748
- Cite this reference work entry

- E. Ronald de Kloet 4 &
- Marian Joëls 5 , 6
155 Accesses
This chapter starts with highlighting the evolution of the stress concept and the discovery of mediators that coordinate stress adaptation. Next, progress in the unraveling of the mechanism underlying the action of these stress mediators is discussed, focusing on glucocorticoids as the end product of the hypothalamus-pituitary-adrenal (HPA) axis. This action exerted by the glucocorticoids is mediated by a dual receptor system: mineralocorticoid (MR) and glucocorticoid receptors (GR). With these receptors as leading theme we present five highlights that illustrate the serendipitous nature of stress research. These five highlights are integrated in the final section which culminates in reflections on the role of stress in mental health. In these reflections we merge the mind-boggling complexity of molecular signaling pathways with neuroendocrine communication, integrating body and brain functions. The new insights will be used during the next decennium to target, in an individual-specific fashion, the stress system with the objective to enhance the quality of life.
This is a preview of subscription content, log in via an institution to check access.
Access this chapter
- Available as PDF
- Read on any device
- Instant download
- Own it forever
- Available as EPUB and PDF
- Durable hardcover edition
- Dispatched in 3 to 5 business days
- Free shipping worldwide - see info
Tax calculation will be finalised at checkout
Purchases are for personal use only
Institutional subscriptions
Abbreviations
5-Hydroxytryptamine = serotonin
Serotonin transporter
Adrenocorticotropic hormone
Adrenalectomy
Apomorphine-susceptible
Corticosterone
Basolateral amygdala
Corticotropin releasing hormone
Dexamethasone
Extracellular regulated kinase 1/2
Glucocorticoid receptor
Hypothalamic-Pituitary-Adrenal axis
Long-term potentiation
Multidrug resistance
Miniature excitatory postsynaptic current
Mineralocorticoid receptor
Pro-opiomelanocortin
Prepulse inhibition
Paraventricular nucleus
Stress hyporesponsive period
Single nucleotide polymorphism
Bale TL, Epperson CN (2017) Sex as a biological variable: who, what, when, why, and how. Neuropsychopharmacology 42:386–396. https://doi.org/10.1038/npp.2016.215 . Epub 2016 Sep 23. PMID: 27658485; PMCID: PMC5399243
Article CAS PubMed Google Scholar
Belsky J, Jonassaint C, Pluess M, Stanton M, Brummett B, Williams R (2009) Vulnerability genes or plasticity genes? Mol Psychiatry 14:746–754
Article CAS Google Scholar
Bonapersona V, Kentrop J, Van Lissa CJ, van der Veen R, Joëls M, Sarabdjitsingh RA (2019) The behavioral phenotype of early life adversity: a 3-level meta-analysis of rodent studies. Neurosci Biobehav Rev 102:299–307. https://doi.org/10.1016/j.neubiorev.2019.04.021 . Epub 2019 Apr 29. PMID: 31047892
Champagne DL, Bagot RC, van Hasselt F, Ramakers G, Meaney MJ, de Kloet ER, Joëls M, Krugers H (2008) Maternal care and hippocampal plasticity: evidence for experience-dependent structural plasticity, altered synaptic functioning, and differential responsiveness to glucocorticoids and stress. J Neurosci 28:6037–6045
Dalm S, Karssen AM, Meijer OC, Belanoff JK, de Kloet ER (2019) Resetting the stress system with a mifepristone challenge. Cell Mol Neurobiol 39(4):503–522. https://doi.org/10.1007/s10571-018-0614-5 . Epub 2018 Sep 1. PMID: 30173378; PMCID: PMC6469632
Daskalakis NP, Bagot RC, Parker KJ, Vinkers CH, de Kloet ER (2013) The three-hit concept of vulnerability and resilience: toward understanding adaptation to early-life adversity outcome. Psychoneuroendocrinology 38:1858–1873. https://doi.org/10.1016/j.psyneuen.2013.06.008 . Epub 2013 Jul 7. PMID: 23838101; PMCID: PMC3773020
Article PubMed PubMed Central Google Scholar
Datson NA, van den Oever JM, Korobko OB, Magarinos AM, de Kloet ER, McEwen BS (2013) Previous history of chronic stress changes the transcriptional response to glucocorticoid challenge in the dentate gyrus region of the male rat hippocampus. Endocrinology 154(9):3261–3272. https://doi.org/10.1210/en.2012-2233 . Epub 2013 Apr 30. PMID: 23633533; PMCID: PMC3749472
Article CAS PubMed PubMed Central Google Scholar
de Kloet ER, Oitzl MS (2003) Who cares for a stressed brain? The mother, the kid or both? Neurobiol Aging 24(Suppl 1):S61–S65; discussion S67–8. PMID: 12829110. https://doi.org/10.1016/s0197-4580(03)00057-5
Article PubMed Google Scholar
De Kloet R, Wallach G, McEwen BS (1975) Differences in corticosterone and dexamethasone binding to rat brain and pituitary. Endocrinology 96:598–609. https://doi.org/10.1210/endo-96-3-598 . PMID: 163728
de Kloet ER, Oitzl MS, Joëls M (1999) Stress and cognition: are corticosteroids good or bad guys? Trends Neurosci 22:422–426
Article Google Scholar
de Kloet ER, Joëls M, Holsboer F (2005) Stress and the brain: from adaptation to disease. Nat Rev Neurosci 6:463–475
de Kloet ER, Meijer OC, de Nicola AF, de Rijk RH, Joëls M (2018) Importance of the brain corticosteroid receptor balance in metaplasticity, cognitive performance and neuro-inflammation. Front Neuroendocrinol 49:124–145. https://doi.org/10.1016/j.yfrne.2018.02.003 . Epub 2018 Feb 8. PMID: 29428549
de Kloet ER, de Kloet SF, de Kloet CS, de Kloet AD (2019) Top-down and bottom-up control of stress-coping. J Neuroendocrinol 31:e12675. https://doi.org/10.1111/jne.12675 . Epub 2019 Feb 1. PMID: 30578574; PMCID: PMC6519262
de Quervain D, Schwabe L, Roozendaal B (2017) Stress, glucocorticoids and memory: implications for treating fear-related disorders. Nat Rev Neurosci 18(1):7–19. https://doi.org/10.1038/nrn.2016.155 . Epub 2016 Nov 24. PMID: 27881856
Derijk RH, van Leeuwen N, Klok MD, Zitman FG (2008) Corticosteroid receptor-gene variants: modulators of the stress-response and implications for mental health. Eur J Pharmacol 585:492–501
Douma EH, de Kloet ER (2020) Stress-induced plasticity and functioning of ventral tegmental dopamine neurons. Neurosci Biobehav Rev 108:48–77. https://doi.org/10.1016/j.neubiorev.2019.10.015 . Epub 2019 Oct 27. PMID: 31666179
Duma D, Collins JB, Chou JW, Cidlowski JA (2010) Sexually dimorphic actions of glucocorticoids provide a link to inflammatory diseases with gender differences in prevalence. Sci Signal 3(143):ra74. https://doi.org/10.1126/scisignal.2001077 . Erratum in: Sci Signal. 2010;3(145):er10. PMID: 20940427; PMCID: PMC4772900
Gasparini S, Resch JM, Narayan SV, Peltekian L, Iverson GN, Karthik S, Geerling JC (2019) Aldosterone-sensitive HSD2 neurons in mice. Brain Struct Funct 224:387–417. https://doi.org/10.1007/s00429-018-1778-y . Epub 2018 Oct 20. PMID: 30343334; PMCID: PMC6369013
Harris AP, Holmes MC, de Kloet ER, Chapman KE, Seckl JR (2013) Mineralocorticoid and glucocorticoid receptor balance in control of HPA axis and behaviour. Psychoneuroendocrinology 38(5):648–658. https://doi.org/10.1016/j.psyneuen.2012.08.007 . Epub 2012 Sep 11. PMID: 22980941
Herman JP, Nawreen N, Smail MA, Cotella EM (2020) Brain mechanisms of HPA axis regulation: neurocircuitry and feedback in context Richard Kvetnansky lecture. Stress 23(6):617–632. https://doi.org/10.1080/10253890.2020.1859475 . Epub 2020 Dec 21. PMID: 33345670; PMCID: PMC8034599.
Hermans EJ, Henckens MJ, Joëls M, Fernández G (2014) Dynamic adaptation of large-scale brain networks in response to acute stressors. Trends Neurosci 37(6):304–14. https://doi.org/10.1016/j.tins.2014.03.006 . Epub 2014 Apr 21. PMID: 24766931.
Henkin RI, Daly RL (1968) Auditory detection and perception in normal man and in patients with adrenal cortical insufficiency: effect of adrenal cortical steroids. J Clin Invest 47:1269–1280. https://doi.org/10.1172/JCI105819 . PMID: 4298249; PMCID: PMC297283
Holsboer F, Ising M (2010) Stress hormone regulation: biological role and translation into therapy. Annu Rev Psychol 61(81–109):C1–C11
Google Scholar
Ingle DJ (1938) Discovery of feedback. Am J Phys 124:369–371
Joëls M (2006) Corticosteroid effects in the brain: U-shape it. Trends Pharmacol Sci 27:244–250
Joëls M, Baram TZ (2009) The neuro-symphony of stress. Nat Rev Neurosci 10:459–466
Joëls M, Pu Z, Wiegert O, Oitzl MS, Krugers HJ (2006) Learning under stress: how does it work? Trends Cogn Sci 10:152–158
Joëls M, Karst H, deRijk R, de Kloet ER (2008) The coming out of the brain mineralocorticoid receptor. Trends Neurosci 31:1–7
Joëls M, Sarabdjitsingh RA, Karst H (2012) Unraveling the time domains of corticosteroid hormone influences on brain activity: rapid, slow, and chronic modes. Pharmacol Rev 64:901–938. https://doi.org/10.1124/pr.112.005892
Joëls M, Karst H, Sarabdjitsingh RA (2018) The stressed brain of humans and rodents. Acta Physiol (Oxf) 223:e13066. https://doi.org/10.1111/apha.13066 . Epub 2018 Apr 16. PMID: 29575542; PMCID: PMC5969253
Karssen AM, Meijer OC, Berry A, Sanjuan Piñol R, de Kloet ER (2005) Low doses of dexamethasone can produce a hypocorticosteroid state in the brain. Endocrinology 146:5587–5595. https://doi.org/10.1210/en.2005-0501 . Epub 2005 Sep 8. PMID: 16150912
Karst H, Joëls M (2016) Severe stress hormone conditions cause an extended window of excitability in the mouse basolateral amygdala. Neuropharmacology 110:175–180. https://doi.org/10.1016/j.neuropharm.2016.07.027 . Epub 2016 Jul 25. PMID: 27460963
Karst H, Berger S, Turiault M, Tronche F, Schütz G, Joëls M (2005) Mineralocorticoid receptors are indispensible for nongenomic modulation of hippocampal glutamate transmission by cortiocsterone. Proc Natl Acad Sci USA 102:19204–19207
Karst H, Berger S, Erdmann G, Schütz G, Joëls M (2010) Metaplasticity of amygdalar responses to the stress hormone corticosterone. Proc Natl Acad Sci USA 107:14449–14454
Klok MD, Giltay EJ, Van der Does AJW, Geleijnse JM, Antypa N, Penninx BWJH, de Geus EJC, Willemsen G, Boomsma DI, Van Leeuwen N, Zitman FG, de Kloet ER, de Rijk RH (2011) A common and functional mineralocorticoid receptor haplotype enhances optimism and protects against depression in females. Transl Psychiatry 1:e62
Krugers HJ, Hoogenraad CC, Groc L (2010) Stress hormones and AMPA receptor trafficking in synaptic plasticity and memory. Nat Rev Neurosci 11:675–681
Levine S (2005) Developmental determinants of sensitivity and resistance to stress. Psychoneuroendocrinology 30:939–994
Lightman SL, Birnie MT, Conway-Campbell BL (2020) Dynamics of ACTH and cortisol secretion and implications for disease. Endocr Rev 41:470–490. https://doi.org/10.1210/endrev/bnaa002 . PMID: 32060528; PMCID: PMC7240781
Lingg RT, Johnson SB, Emmons EB, Anderson RM, Romig-Martin SA, Narayanan NS, McGaugh JL, LaLumiere RT, Radley JJ (2020) Bed nuclei of the stria terminalis modulate memory consolidation via glucocorticoid-dependent and -independent circuits. Proc Natl Acad Sci U S A 117(14):8104–8114. https://doi.org/10.1073/pnas.1915501117 . Epub 2020 Mar 19. PMID: 32193346; PMCID: PMC7148582.
Lupien SJ, McEwen BS, Gunnar MR, Heim C (2009) Effects of stress throughout the lifespan on the brain, behaviour and cognition. Nat Rev Neurosci 10:434–445
McEwen BS, Wingfield JC (2010) What is in a name? Integrating homeostasis, allostasis and stress. Horm Behav 57:105–111
McEwen BS, Weiss JM, Schwartz LS (1968) Selective retention of corticosterone by limbic structures in rat brain. Nature 220:911–912. https://doi.org/10.1038/220911a0 . PMID: 4301849
Meijer OC, de Lange EC, Breimer DD, de Boer AG, Workel JO, de Kloet ER (1998) Penetration of dexamethasone into brain glucocorticoid targets is enhanced in mdr1A P-glycoprotein knockout mice. Endocrinology 139:1789–1793. https://doi.org/10.1210/endo.139.4.5917 . PMID: 9528963
Molendijk ML, de Kloet ER (2021) Forced swim stressor: trends in usage and mechanistic consideration. Eur J Neurosci. https://doi.org/10.1111/ejn.15139 . Epub ahead of print. PMID: 33548153
Munck A, Guyre PM, Holbrook NJ (1984) Physiological functions of glucocorticoids in stress and their relation to pharmacological actions. Endocr. Rev. 5, 25–44. https://doi.org/10.1210/edrv-5-1-25 .
Oakley RH, Cidlowski JA (2011) Cellular processing of the glucocorticoid receptor gene and protein: new mechanisms for generating tissue-specific actions of glucocorticoids. J Biol Chem 286:3177–3184
Obleser J, Kreitewolf J, Vielhauer R, Lindner F, David C, Oster H, Tune S (2021) Circadian fluctuations in glucocorticoid level predict perceptual discrimination sensitivity. iScience. 24(4):102345. https://doi.org/10.1016/j.isci.2021.102345 . PMID: 33870139; PMCID: PMC8047178.
Oitzl MS, de Kloet ER (1992) Selective corticosteroid antagonists modulate specific aspects of spatial orientation learning. Behav Neurosci 106:62–71
Oitzl MS, Champagne DL, van der Veen R, de Kloet ER (2010) Brain development under stress: hypotheses of glucocorticoid actions revisited. Neurosci Biobehav Rev 34:853–866
Oitzl MS, Schwabe L, Aggleton JP (2012) Memory formation: its changing face. Neurosci Biobehav Rev. [Epub ahead of print]
Picard M, Sandi C (2021) The social nature of mitochondria: implications for human health. Neurosci Biobehav Rev 120:595–610. https://doi.org/10.1016/j.neubiorev.2020.04.017 . Epub 2020 Jul 8. PMID: 32651001
Radley JJ, Johnson SB (2018) Anteroventral bed nuclei of the stria terminalis neurocircuitry: towards an integration of HPA axis modulation with coping behaviors – Curt Richter Award Paper 2017. Psychoneuroendocrinology 89:239–249. https://doi.org/10.1016/j.psyneuen.2017.12.005 . Epub 2017 Dec 24. PMID: 29395488; PMCID: PMC5878723
Reul JM, de Kloet ER (1985) Two receptor systems for corticosterone in rat brain: microdistribution and differential occupation. Endocrinology 117:2505–2511. https://doi.org/10.1210/endo-117-6-2505 . PMID: 2998738
Roozendaal B, McEwen BS, Chattarji S (2009) Stress, memory and the amygdala. Nat Rev Neurosci 10:423–433
Sarabdjitsingh RA, Isenia S, Polman A, Mijalkovic J, Lachize S, Datson N, de Kloet ER, Meijer OC (2010) Disrupted corticosterone pulsatile patterns attenuate responsiveness to glucocorticoid signaling in rat brain. Endocrinology 151:1177–1186
Sibug RM, de Koning J, Tijssen AM, de Ruiter MC, de Kloet ER, Helmerhorst FM (2005) Urinary gonadotrophins but not recombinant gonadotrophins reduce expression of VEGF120 and its receptors flt-1 and flk-1 in the mouse uterus during the peri-implantation period. Hum Reprod 20:649–656
Spiga F, Zavala E, Walker JJ, Zhao Z, Terry JR, Lightman SL (2017) Dynamic responses of the adrenal steroidogenic regulatory network. Proc Natl Acad Sci USA 114:E6466–E6474. https://doi.org/10.1073/pnas.1703779114 . Epub 2017 Jul 17. PMID: 28716938; PMCID: PMC5547620
Tausk M, 1952. Hat die Nebennierrinde tatsächlich eine Verteidigungsfunktion? Das Horm. 3
Taylor SE, Klein LC, Lewis BP, Gruenewald TL, Gurung RA, Updegraff JA (2000) Biobehavioral responses to stress in females: tend-and-befriend, not fight-or-flight. Psychol Rev 107(3):411–429. https://doi.org/10.1037/0033-295x.107.3.411 . PMID: 10941275
Zorn JV, Schür RR, Boks MP, Kahn RS, Joëls M, Vinkers CH (2017) Cortisol stress reactivity across psychiatric disorders: a systematic review and meta-analysis. Psychoneuroendocrinology 77:25–36. https://doi.org/10.1016/j.psyneuen.2016.11.036 . Epub 2016 Dec 8. PMID: 28012291
Download references
Author information
Authors and affiliations.
Department of Medicine, Division of Endocrinology, Leiden University Medical Center, Leiden, The Netherlands
E. Ronald de Kloet
Department of Neuroscience and Pharmacology, University Medical Center Utrecht Rudolf Magnus Institute of Neuroscience, Utrecht, The Netherlands
Marian Joëls
University of Groningen, University Medical Center Groningen, Groningen, The Netherlands
You can also search for this author in PubMed Google Scholar
Corresponding author
Correspondence to E. Ronald de Kloet .
Editor information
Editors and affiliations.
Laboratory of Neurobiology and Behavior, The Rockefeller University, New York, NY, USA
Donald W. Pfaff
National Institute on Drug Abuse, Rockville, MD, USA
Nora D. Volkow
Department of Psychiatry, University of California, San Francisco, San Francisco, CA, USA
John L. Rubenstein
Rights and permissions
Reprints and permissions
Copyright information
© 2022 Springer Science+Business Media, LLC, part of Springer Nature
About this entry
Cite this entry.
de Kloet, E.R., Joëls, M. (2022). Stress Research: Past, Present, and Future. In: Pfaff, D.W., Volkow, N.D., Rubenstein, J.L. (eds) Neuroscience in the 21st Century. Springer, Cham. https://doi.org/10.1007/978-3-030-88832-9_72
Download citation
DOI : https://doi.org/10.1007/978-3-030-88832-9_72
Published : 18 October 2022
Publisher Name : Springer, Cham
Print ISBN : 978-3-030-88831-2
Online ISBN : 978-3-030-88832-9
eBook Packages : Biomedical and Life Sciences Reference Module Biomedical and Life Sciences
Share this entry
Anyone you share the following link with will be able to read this content:
Sorry, a shareable link is not currently available for this article.
Provided by the Springer Nature SharedIt content-sharing initiative
- Publish with us
Policies and ethics
- Find a journal
- Track your research
- Tools and Resources
- Customer Services
- Affective Science
- Biological Foundations of Psychology
- Clinical Psychology: Disorders and Therapies
- Cognitive Psychology/Neuroscience
- Developmental Psychology
- Educational/School Psychology
- Forensic Psychology
- Health Psychology
- History and Systems of Psychology
- Individual Differences
- Methods and Approaches in Psychology
- Neuropsychology
- Organizational and Institutional Psychology
- Personality
- Psychology and Other Disciplines
- Social Psychology
- Sports Psychology
- Share This Facebook LinkedIn Twitter
Article contents
Work, stress, coping, and stress management.
- Sharon Glazer Sharon Glazer University of Baltimore
- and Cong Liu Cong Liu Hofstra University
- https://doi.org/10.1093/acrefore/9780190236557.013.30
- Published online: 26 April 2017
Work stress refers to the process of job stressors, or stimuli in the workplace, leading to strains, or negative responses or reactions. Organizational development refers to a process in which problems or opportunities in the work environment are identified, plans are made to remediate or capitalize on the stimuli, action is taken, and subsequently the results of the plans and actions are evaluated. When organizational development strategies are used to assess work stress in the workplace, the actions employed are various stress management interventions. Two key factors tying work stress and organizational development are the role of the person and the role of the environment. In order to cope with work-related stressors and manage strains, organizations must be able to identify and differentiate between factors in the environment that are potential sources of stressors and how individuals perceive those factors. Primary stress management interventions focus on preventing stressors from even presenting, such as by clearly articulating workers’ roles and providing necessary resources for employees to perform their job. Secondary stress management interventions focus on a person’s appraisal of job stressors as a threat or challenge, and the person’s ability to cope with the stressors (presuming sufficient internal resources, such as a sense of meaningfulness in life, or external resources, such as social support from a supervisor). When coping is not successful, strains may develop. Tertiary stress management interventions attempt to remediate strains, by addressing the consequence itself (e.g., diabetes management) and/or the source of the strain (e.g., reducing workload). The person and/or the organization may be the targets of the intervention. The ultimate goal of stress management interventions is to minimize problems in the work environment, intensify aspects of the work environment that create a sense of a quality work context, enable people to cope with stressors that might arise, and provide tools for employees and organizations to manage strains that might develop despite all best efforts to create a healthy workplace.
- stress management
- organization development
- organizational interventions
- stress theories and frameworks
Introduction
Work stress is a generic term that refers to work-related stimuli (aka job stressors) that may lead to physical, behavioral, or psychological consequences (i.e., strains) that affect both the health and well-being of the employee and the organization. Not all stressors lead to strains, but all strains are a result of stressors, actual or perceived. Common terms often used interchangeably with work stress are occupational stress, job stress, and work-related stress. Terms used interchangeably with job stressors include work stressors, and as the specificity of the type of stressor might include psychosocial stressor (referring to the psychological experience of work demands that have a social component, e.g., conflict between two people; Hauke, Flintrop, Brun, & Rugulies, 2011 ), hindrance stressor (i.e., a stressor that prevents goal attainment; Cavanaugh, Boswell, Roehling, & Boudreau, 2000 ), and challenge stressor (i.e., a stressor that is difficult, but attainable and possibly rewarding to attain; Cavanaugh et al., 2000 ).
Stress in the workplace continues to be a highly pervasive problem, having both direct negative effects on individuals experiencing it and companies paying for it, and indirect costs vis à vis lost productivity (Dopkeen & DuBois, 2014 ). For example, U.K. public civil servants’ work-related stress rose from 10.8% in 2006 to 22.4% in 2013 and about one-third of the workforce has taken more than 20 days of leave due to stress-related ill-health, while well over 50% are present at work when ill (French, 2015 ). These findings are consistent with a report by the International Labor Organization (ILO, 2012 ), whereby 50% to 60% of all workdays are lost due to absence attributed to factors associated with work stress.
The prevalence of work-related stress is not diminishing despite improvements in technology and employment rates. The sources of stress, such as workload, seem to exacerbate with improvements in technology (Coovert & Thompson, 2003 ). Moreover, accessibility through mobile technology and virtual computer terminals is linking people to their work more than ever before (ILO, 2012 ; Tarafdar, Tu, Ragu-Nathan, & Ragu-Nathan, 2007 ). Evidence of this kind of mobility and flexibility is further reinforced in a June 2007 survey of 4,025 email users (over 13 years of age); AOL reported that four in ten survey respondents reported planning their vacations around email accessibility and 83% checked their emails at least once a day while away (McMahon, 2007 ). Ironically, despite these mounting work-related stressors and clear financial and performance outcomes, some individuals are reporting they are less “stressed,” but only because “stress has become the new normal” (Jayson, 2012 , para. 4).
This new normal is likely the source of psychological and physiological illness. Siegrist ( 2010 ) contends that conditions in the workplace, particularly psychosocial stressors that are perceived as unfavorable relationships with others and self, and an increasingly sedentary lifestyle (reinforced with desk jobs) are increasingly contributing to cardiovascular disease. These factors together justify a need to continue on the path of helping individuals recognize and cope with deleterious stressors in the work environment and, equally important, to find ways to help organizations prevent harmful stressors over which they have control, as well as implement policies or mechanisms to help employees deal with these stressors and subsequent strains. Along with a greater focus on mitigating environmental constraints are interventions that can be used to prevent anxiety, poor attitudes toward the workplace conditions and arrangements, and subsequent cardiovascular illness, absenteeism, and poor job performance (Siegrist, 2010 ).
Even the ILO has presented guidance on how the workplace can help prevent harmful job stressors (aka hindrance stressors) or at least help workers cope with them. Consistent with the view that well-being is not the absence of stressors or strains and with the view that positive psychology offers a lens for proactively preventing stressors, the ILO promotes increasing preventative risk assessments, interventions to prevent and control stressors, transparent organizational communication, worker involvement in decision-making, networks and mechanisms for workplace social support, awareness of how working and living conditions interact, safety, health, and well-being in the organization (ILO, n.d. ). The field of industrial and organizational (IO) psychology supports the ILO’s recommendations.
IO psychology views work stress as the process of a person’s interaction with multiple aspects of the work environment, job design, and work conditions in the organization. Interventions to manage work stress, therefore, focus on the psychosocial factors of the person and his or her relationships with others and the socio-technical factors related to the work environment and work processes. Viewing work stress from the lens of the person and the environment stems from Kurt Lewin’s ( 1936 ) work that stipulates a person’s state of mental health and behaviors are a function of the person within a specific environment or situation. Aspects of the work environment that affect individuals’ mental states and behaviors include organizational hierarchy, organizational climate (including processes, policies, practices, and reward structures), resources to support a person’s ability to fulfill job duties, and management structure (including leadership). Job design refers to each contributor’s tasks and responsibilities for fulfilling goals associated with the work role. Finally, working conditions refers not only to the physical environment, but also the interpersonal relationships with other contributors.
Each of the conditions that are identified in the work environment may be perceived as potentially harmful or a threat to the person or as an opportunity. When a stressor is perceived as a threat to attaining desired goals or outcomes, the stressor may be labeled as a hindrance stressor (e.g., LePine, Podsakoff, & Lepine, 2005 ). When the stressor is perceived as an opportunity to attain a desired goal or end state, it may be labeled as a challenge stressor. According to LePine and colleagues’ ( 2005 ), both challenge (e.g., time urgency, workload) and hindrance (e.g., hassles, role ambiguity, role conflict) stressors could lead to strains (as measured by “anxiety, depersonalization, depression, emotional exhaustion, frustration, health complaints, hostility, illness, physical symptoms, and tension” [p. 767]). However, challenge stressors positively relate with motivation and performance, whereas hindrance stressors negatively relate with motivation and performance. Moreover, motivation and strains partially mediate the relationship between hindrance and challenge stressors with performance.
Figure 1. Organizational development frameworks to guide identification of work stress and interventions.
In order to (1) minimize any potential negative effects from stressors, (2) increase coping skills to deal with stressors, or (3) manage strains, organizational practitioners or consultants will devise organizational interventions geared toward prevention, coping, and/or stress management. Ultimately, toxic factors in the work environment can have deleterious effects on a person’s physical and psychological well-being, as well as on an organization’s total health. It behooves management to take stock of the organization’s health, which includes the health and well-being of its employees, if the organization wishes to thrive and be profitable. According to Page and Vella-Brodrick’s ( 2009 ) model of employee well-being, employee well-being results from subjective well-being (i.e., life satisfaction and general positive or negative affect), workplace well-being (composed of job satisfaction and work-specific positive or negative affect), and psychological well-being (e.g., self-acceptance, positive social relations, mastery, purpose in life). Job stressors that become unbearable are likely to negatively affect workplace well-being and thus overall employee well-being. Because work stress is a major organizational pain point and organizations often employ organizational consultants to help identify and remediate pain points, the focus here is on organizational development (OD) frameworks; several work stress frameworks are presented that together signal areas where organizations might focus efforts for change in employee behaviors, attitudes, and performance, as well as the organization’s performance and climate. Work stress, interventions, and several OD and stress frameworks are depicted in Figure 1 .
The goals are: (1) to conceptually define and clarify terms associated with stress and stress management, particularly focusing on organizational factors that contribute to stress and stress management, and (2) to present research that informs current knowledge and practices on workplace stress management strategies. Stressors and strains will be defined, leading OD and work stress frameworks that are used to organize and help organizations make sense of the work environment and the organization’s responsibility in stress management will be explored, and stress management will be explained as an overarching thematic label; an area of study and practice that focuses on prevention (primary) interventions, coping (secondary) interventions, and managing strains (tertiary) interventions; as well as the label typically used to denote tertiary interventions. Suggestions for future research and implications toward becoming a healthy organization are presented.
Defining Stressors and Strains
Work-related stressors or job stressors can lead to different kinds of strains individuals and organizations might experience. Various types of stress management interventions, guided by OD and work stress frameworks, may be employed to prevent or cope with job stressors and manage strains that develop(ed).
A job stressor is a stimulus external to an employee and a result of an employee’s work conditions. Example job stressors include organizational constraints, workplace mistreatments (such as abusive supervision, workplace ostracism, incivility, bullying), role stressors, workload, work-family conflicts, errors or mistakes, examinations and evaluations, and lack of structure (Jex & Beehr, 1991 ; Liu, Spector, & Shi, 2007 ; Narayanan, Menon, & Spector, 1999 ). Although stressors may be categorized as hindrances and challenges, there is not yet sufficient information to be able to propose which stress management interventions would better serve to reduce those hindrance stressors or to reduce strain-producing challenge stressors while reinforcing engagement-producing challenge stressors.
Organizational Constraints
Organizational constraints may be hindrance stressors as they prevent employees from translating their motivation and ability into high-level job performance (Peters & O’Connor, 1980 ). Peters and O’Connor ( 1988 ) defined 11 categories of organizational constraints: (1) job-related information, (2) budgetary support, (3) required support, (4) materials and supplies, (5) required services and help from others, (6) task preparation, (7) time availability, (8) the work environment, (9) scheduling of activities, (10) transportation, and (11) job-relevant authority. The inhibiting effect of organizational constraints may be due to the lack of, inadequacy of, or poor quality of these categories.
Workplace Mistreatment
Workplace mistreatment presents a cluster of interpersonal variables, such as interpersonal conflict, bullying, incivility, and workplace ostracism (Hershcovis, 2011 ; Tepper & Henle, 2011 ). Typical workplace mistreatment behaviors include gossiping, rude comments, showing favoritism, yelling, lying, and ignoring other people at work (Tepper & Henle, 2011 ). These variables relate to employees’ psychological well-being, physical well-being, work attitudes (e.g., job satisfaction and organizational commitment), and turnover intention (e.g., Hershcovis, 2011 ; Spector & Jex, 1998 ). Some researchers differentiated the source of mistreatment, such as mistreatment from one’s supervisor versus mistreatment from one’s coworker (e.g., Bruk-Lee & Spector, 2006 ; Frone, 2000 ; Liu, Liu, Spector, & Shi, 2011 ).
Role Stressors
Role stressors are demands, constraints, or opportunities a person perceives to be associated, and thus expected, with his or her work role(s) across various situations. Three commonly studied role stressors are role ambiguity, role conflict, and role overload (Glazer & Beehr, 2005 ; Kahn, Wolfe, Quinn, Snoek, & Rosenthal, 1964 ). Role ambiguity in the workplace occurs when an employee lacks clarity regarding what performance-related behaviors are expected of him or her. Role conflict refers to situations wherein an employee receives incompatible role requests from the same or different supervisors or the employee is asked to engage in work that impedes his or her performance in other work or nonwork roles or clashes with his or her values. Role overload refers to excessive demands and insufficient time (quantitative) or knowledge (qualitative) to complete the work. The construct is often used interchangeably with workload, though role overload focuses more on perceived expectations from others about one’s workload. These role stressors significantly relate to low job satisfaction, low organizational commitment, low job performance, high tension or anxiety, and high turnover intention (Abramis, 1994 ; Glazer & Beehr, 2005 ; Jackson & Schuler, 1985 ).
Excessive workload is one of the most salient stressors at work (e.g., Liu et al., 2007 ). There are two types of workload: quantitative and qualitative workload (LaRocco, Tetrick, & Meder, 1989 ; Parasuraman & Purohit, 2000 ). Quantitative workload refers to the excessive amount of work one has. In a summary of a Chartered Institute of Personnel & Development Report from 2006 , Dewe and Kompier ( 2008 ) noted that quantitative workload was one of the top three stressors workers experienced at work. Qualitative workload refers to the difficulty of work. Workload also differs by the type of the load. There are mental workload and physical workload (Dwyer & Ganster, 1991 ). Excessive physical workload may result in physical discomfort or illness. Excessive mental workload will cause psychological distress such as anxiety or frustration (Bowling & Kirkendall, 2012 ). Another factor affecting quantitative workload is interruptions (during the workday). Lin, Kain, and Fritz ( 2013 ) found that interruptions delay completion of job tasks, thus adding to the perception of workload.
Work-Family Conflict
Work-family conflict is a form of inter-role conflict in which demands from one’s work domain and one’s family domain are incompatible to some extent (Greenhaus & Beutell, 1985 ). Work can interfere with family (WIF) and/or family can interfere with work (FIW) due to time-related commitments to participating in one domain or another, incompatible behavioral expectations, or when strains in one domain carry over to the other (Greenhaus & Beutell, 1985 ). Work-family conflict significantly relates to work-related outcomes (e.g., job satisfaction, organizational commitment, turnover intention, burnout, absenteeism, job performance, job strains, career satisfaction, and organizational citizenship behaviors), family-related outcomes (e.g., marital satisfaction, family satisfaction, family-related performance, family-related strains), and domain-unspecific outcomes (e.g., life satisfaction, psychological strain, somatic or physical symptoms, depression, substance use or abuse, and anxiety; Amstad, Meier, Fasel, Elfering, & Semmer, 2011 ).
Individuals and organizations can experience work-related strains. Sometimes organizations will experience strains through the employee’s negative attitudes or strains, such as that a worker’s absence might yield lower production rates, which would roll up into an organizational metric of organizational performance. In the industrial and organizational (IO) psychology literature, organizational strains are mostly observed as macro-level indicators, such as health insurance costs, accident-free days, and pervasive problems with company morale. In contrast, individual strains, usually referred to as job strains, are internal to an employee. They are responses to work conditions and relate to health and well-being of employees. In other words, “job strains are adverse reactions employees have to job stressors” (Spector, Chen, & O’Connell, 2000 , p. 211). Job strains tend to fall into three categories: behavioral, physical, and psychological (Jex & Beehr, 1991 ).
Behavioral strains consist of actions that employees take in response to job stressors. Examples of behavioral strains include employees drinking alcohol in the workplace or intentionally calling in sick when they are not ill (Spector et al., 2000 ). Physical strains consist of health symptoms that are physiological in nature that employees contract in response to job stressors. Headaches and ulcers are examples of physical strains. Lastly, psychological strains are emotional reactions and attitudes that employees have in response to job stressors. Examples of psychological strains are job dissatisfaction, anxiety, and frustration (Spector et al., 2000 ). Interestingly, research studies that utilize self-report measures find that most job strains experienced by employees tend to be psychological strains (Spector et al., 2000 ).
Leading Frameworks
Organizations that are keen on identifying organizational pain points and remedying them through organizational campaigns or initiatives often discover the pain points are rooted in work-related stressors and strains and the initiatives have to focus on reducing workers’ stress and increasing a company’s profitability. Through organizational climate surveys, for example, companies discover that aspects of the organization’s environment, including its policies, practices, reward structures, procedures, and processes, as well as employees at all levels of the company, are contributing to the individual and organizational stress. Recent studies have even begun to examine team climates for eustress and distress assessed in terms of team members’ homogenous psychological experience of vigor, efficacy, dedication, and cynicism (e.g., Kożusznik, Rodriguez, & Peiro, 2015 ).
Each of the frameworks presented advances different aspects that need to be identified in order to understand the source and potential remedy for stressors and strains. In some models, the focus is on resources, in others on the interaction of the person and environment, and in still others on the role of the person in the workplace. Few frameworks directly examine the role of the organization, but the organization could use these frameworks to plan interventions that would minimize stressors, cope with existing stressors, and prevent and/or manage strains. One of the leading frameworks in work stress research that is used to guide organizational interventions is the person and environment (P-E) fit (French & Caplan, 1972 ). Its precursor is the University of Michigan Institute for Social Research’s (ISR) role stress model (Kahn, Wolfe, Quinn, Snoek, & Rosenthal, 1964 ) and Lewin’s Field Theory. Several other theories have since evolved from the P-E fit framework, including Karasek and Theorell’s ( 1990 ), Karasek ( 1979 ) Job Demands-Control Model (JD-C), the transactional framework (Lazarus & Folkman, 1984 ), Conservation of Resources (COR) theory (Hobfoll, 1989 ), and Siegrist’s ( 1996 ) Effort-Reward Imbalance (ERI) Model.
Field Theory
The premise of Kahn et al.’s ( 1964 ) role stress theory is Lewin’s ( 1997 ) Field Theory. Lewin purported that behavior and mental events are a dynamic function of the whole person, including a person’s beliefs, values, abilities, needs, thoughts, and feelings, within a given situation (field or environment), as well as the way a person represents his or her understanding of the field and behaves in that space. Lewin explains that work-related strains are a result of individuals’ subjective perceptions of objective factors, such as work roles, relationships with others in the workplace, as well as personality indicators, and can be used to predict people’s reactions, including illness. Thus, to make changes to an organizational system, it is necessary to understand a field and try to move that field from the current state to the desired state. Making this move necessitates identifying mechanisms influencing individuals.
Role Stress Theory
Role stress theory mostly isolates the perspective a person has about his or her work-related responsibilities and expectations to determine how those perceptions relate with a person’s work-related strains. However, those relationships have been met with somewhat varied results, which Glazer and Beehr ( 2005 ) concluded might be a function of differences in culture, an environmental factor often neglected in research. Kahn et al.’s ( 1964 ) role stress theory, coupled with Lewin’s ( 1936 ) Field Theory, serves as the foundation for the P-E fit theory. Lewin ( 1936 ) wrote, “Every psychological event depends upon the state of the person and at the same time on the environment” (p. 12). Researchers of IO psychology have narrowed the environment to the organization or work team. This narrowed view of the organizational environment is evident in French and Caplan’s ( 1972 ) P-E fit framework.
Person-Environment Fit Theory
The P-E fit framework focuses on the extent to which there is congruence between the person and a given environment, such as the organization (Caplan, 1987 ; Edwards, 2008 ). For example, does the person have the necessary skills and abilities to fulfill an organization’s demands, or does the environment support a person’s desire for autonomy (i.e., do the values align?) or fulfill a person’s needs (i.e., a person’s needs are rewarded). Theoretically and empirically, the greater the person-organization fit, the greater a person’s job satisfaction and organizational commitment, the less a person’s turnover intention and work-related stress (see meta-analyses by Assouline & Meir, 1987 ; Kristof-Brown, Zimmerman, & Johnson, 2005 ; Verquer, Beehr, & Wagner, 2003 ).
Job Demands-Control/Support (JD-C/S) and Job Demands-Resources (JD-R) Model
Focusing more closely on concrete aspects of work demands and the extent to which a person perceives he or she has control or decision latitude over those demands, Karasek ( 1979 ) developed the JD-C model. Karasek and Theorell ( 1990 ) posited that high job demands under conditions of little decision latitude or control yield high strains, which have varied implications on the health of an organization (e.g., in terms of high turnover, employee ill-health, poor organizational performance). This theory was modified slightly to address not only control, but also other resources that could protect a person from unruly job demands, including support (aka JD-C/S, Johnson & Hall, 1988 ; and JD-R, Bakker, van Veldhoven, & Xanthopoulou, 2010 ). Whether focusing on control or resources, both they and job demands are said to reflect workplace characteristics, while control and resources also represent coping strategies or tools (Siegrist, 2010 ).
Despite the glut of research testing the JD-C and JD-R, results are somewhat mixed. Testing the interaction between job demands and control, Beehr, Glaser, Canali, and Wallwey ( 2001 ) did not find empirical support for the JD-C theory. However, Dawson, O’Brien, and Beehr ( 2016 ) found that high control and high support buffered against the independent deleterious effects of interpersonal conflict, role conflict, and organizational politics (demands that were categorized as hindrance stressors) on anxiety, as well as the effects of interpersonal conflict and organizational politics on physiological symptoms, but control and support did not moderate the effects between challenge stressors and strains. Coupled with Bakker, Demerouti, and Sanz-Vergel’s ( 2014 ) note that excessive job demands are a source of strain, but increased job resources are a source of engagement, Dawson et al.’s results suggest that when an organization identifies that demands are hindrances, it can create strategies for primary (preventative) stress management interventions and attempt to remove or reduce such work demands. If the demands are challenging, though manageable, but latitude to control the challenging stressors and support are insufficient, the organization could modify practices and train employees on adopting better strategies for meeting or coping (secondary stress management intervention) with the demands. Finally, if the organization can neither afford to modify the demands or the level of control and support, it will be necessary for the organization to develop stress management (tertiary) interventions to deal with the inevitable strains.
Conservation of Resources Theory
The idea that job resources reinforce engagement in work has been propagated in Hobfoll’s ( 1989 ) Conservation of Resources (COR) theory. COR theory also draws on the foundational premise that people’s mental health is a function of the person and the environment, forwarding that how people interpret their environment (including the societal context) affects their stress levels. Hobfoll focuses on resources such as objects, personal characteristics, conditions, or energies as particularly instrumental to minimizing strains. He asserts that people do whatever they can to protect their valued resources. Thus, strains develop when resources are threatened to be taken away, actually taken away, or when additional resources are not attainable after investing in the possibility of gaining more resources (Hobfoll, 2001 ). By extension, organizations can invest in activities that would minimize resource loss and create opportunities for resource gains and thus have direct implications for devising primary and secondary stress management interventions.
Transactional Framework
Lazarus and Folkman ( 1984 ) developed the widely studied transactional framework of stress. This framework holds as a key component the cognitive appraisal process. When individuals perceive factors in the work environment as a threat (i.e., primary appraisal), they will scan the available resources (external or internal to himself or herself) to cope with the stressors (i.e., secondary appraisal). If the coping resources provide minimal relief, strains develop. Until recently, little attention has been given to the cognitive appraisal associated with different work stressors (Dewe & Kompier, 2008 ; Liu & Li, 2017 ). In a study of Polish and Spanish social care service providers, stressors appraised as a threat related positively to burnout and less engagement, but stressors perceived as challenges yielded greater engagement and less burnout (Kożusznik, Rodriguez, & Peiro, 2012 ). Similarly, Dawson et al. ( 2016 ) found that even with support and control resources, hindrance demands were more strain-producing than challenge demands, suggesting that appraisal of the stressor is important. In fact, “many people respond well to challenging work” (Beehr et al., 2001 , p. 126). Kożusznik et al. ( 2012 ) recommend training employees to change the way they view work demands in order to increase engagement, considering that part of the problem may be about how the person appraises his or her environment and, thus, copes with the stressors.
Effort-Reward Imbalance
Siegrist’s ( 1996 ) Model of Effort-Reward Imbalance (ERI) focuses on the notion of social reciprocity, such that a person fulfills required work tasks in exchange for desired rewards (Siegrist, 2010 ). ERI sheds light on how an imbalance in a person’s expectations of an organization’s rewards (e.g., pay, bonus, sense of advancement and development, job security) in exchange for a person’s efforts, that is a break in one’s work contract, leads to negative responses, including long-term ill-health (Siegrist, 2010 ; Siegrist et al., 2014 ). In fact, prolonged perception of a work contract imbalance leads to adverse health, including immunological problems and inflammation, which contribute to cardiovascular disease (Siegrist, 2010 ). The model resembles the relational and interactional psychological contract theory in that it describes an employee’s perception of the terms of the relationship between the person and the workplace, including expectations of performance, job security, training and development opportunities, career progression, salary, and bonuses (Thomas, Au, & Ravlin, 2003 ). The psychological contract, like the ERI model, focuses on social exchange. Furthermore, the psychological contract, like stress theories, are influenced by cultural factors that shape how people interpret their environments (Glazer, 2008 ; Thomas et al., 2003 ). Violations of the psychological contract will negatively affect a person’s attitudes toward the workplace and subsequent health and well-being (Siegrist, 2010 ). To remediate strain, Siegrist ( 2010 ) focuses on both the person and the environment, recognizing that the organization is particularly responsible for changing unfavorable work conditions and the person is responsible for modifying his or her reactions to such conditions.
Stress Management Interventions: Primary, Secondary, and Tertiary
Remediation of work stress and organizational development interventions are about realigning the employee’s experiences in the workplace with factors in the environment, as well as closing the gap between the current environment and the desired environment. Work stress develops when an employee perceives the work demands to exceed the person’s resources to cope and thus threatens employee well-being (Dewe & Kompier, 2008 ). Likewise, an organization’s need to change arises when forces in the environment are creating a need to change in order to survive (see Figure 1 ). Lewin’s ( 1951 ) Force Field Analysis, the foundations of which are in Field Theory, is one of the first organizational development intervention tools presented in the social science literature. The concept behind Force Field Analysis is that in order to survive, organizations must adapt to environmental forces driving a need for organizational change and remove restraining forces that create obstacles to organizational change. In order to do this, management needs to delineate the current field in which the organization is functioning, understand the driving forces for change, identify and dampen or eliminate the restraining forces against change. Several models for analyses may be applied, but most approaches are variations of organizational climate surveys.
Through organizational surveys, workers provide management with a snapshot view of how they perceive aspects of their work environment. Thus, the view of the health of an organization is a function of several factors, chief among them employees’ views (i.e., the climate) about the workplace (Lewin, 1951 ). Indeed, French and Kahn ( 1962 ) posited that well-being depends on the extent to which properties of the person and properties of the environment align in terms of what a person requires and the resources available in a given environment. Therefore, only when properties of the person and properties of the environment are sufficiently understood can plans for change be developed and implemented targeting the environment (e.g., change reporting structures to relieve, and thus prevent future, communication stressors) and/or the person (e.g., providing more autonomy, vacation days, training on new technology). In short, climate survey findings can guide consultants about the emphasis for organizational interventions: before a problem arises aka stress prevention, e.g., carefully crafting job roles), when a problem is present, but steps are taken to mitigate their consequences (aka coping, e.g., providing social support groups), and/or once strains develop (aka. stress management, e.g., healthcare management policies).
For each of the primary (prevention), secondary (coping), and tertiary (stress management) techniques the target for intervention can be the entire workforce, a subset of the workforce, or a specific person. Interventions that target the entire workforce may be considered organizational interventions, as they have direct implications on the health of all individuals and consequently the health of the organization. Several interventions categorized as primary and secondary interventions may also be implemented after strains have developed and after it has been discerned that a person or the organization did not do enough to mitigate stressors or strains (see Figure 1 ). The designation of many of the interventions as belonging to one category or another may be viewed as merely a suggestion.
Primary Interventions (Preventative Stress Management)
Before individuals begin to perceive work-related stressors, organizations engage in stress prevention strategies, such as providing people with resources (e.g., computers, printers, desk space, information about the job role, organizational reporting structures) to do their jobs. However, sometimes the institutional structures and resources are insufficient or ambiguous. Scholars and practitioners have identified several preventative stress management strategies that may be implemented.
Planning and Time Management
When employees feel quantitatively overloaded, sometimes the remedy is improving the employees’ abilities to plan and manage their time (Quick, Quick, Nelson, & Hurrell, 2003 ). Planning is a future-oriented activity that focuses on conceptual and comprehensive work goals. Time management is a behavior that focuses on organizing, prioritizing, and scheduling work activities to achieve short-term goals. Given the purpose of time management, it is considered a primary intervention, as engaging in time management helps to prevent work tasks from mounting and becoming unmanageable, which would subsequently lead to adverse outcomes. Time management comprises three fundamental components: (1) establishing goals, (2) identifying and prioritizing tasks to fulfill the goals, and (3) scheduling and monitoring progress toward goal achievement (Peeters & Rutte, 2005 ). Workers who employ time management have less role ambiguity (Macan, Shahani, Dipboye, & Philips, 1990 ), psychological stress or strain (Adams & Jex, 1999 ; Jex & Elaqua, 1999 ; Macan et al., 1990 ), and greater job satisfaction (Macan, 1994 ). However, Macan ( 1994 ) did not find a relationship between time management and performance. Still, Claessens, van Eerde, Rutte, and Roe ( 2004 ) found that perceived control of time partially mediated the relationships between planning behavior (an indicator of time management), job autonomy, and workload on one hand, and job strains, job satisfaction, and job performance on the other hand. Moreover, Peeters and Rutte ( 2005 ) observed that teachers with high work demands and low autonomy experienced more burnout when they had poor time management skills.
Person-Organization Fit
Just as it is important for organizations to find the right person for the job and organization, so is it the responsibility of a person to choose to work at the right organization—an organization that fulfills the person’s needs and upholds the values important to the individual, as much as the person fulfills the organization’s needs and adapts to its values. When people fit their employing organizations they are setting themselves up for experiencing less strain-producing stressors (Kristof-Brown et al., 2005 ). In a meta-analysis of 62 person-job fit studies and 110 person-organization fit studies, Kristof-Brown et al. ( 2005 ) found that person-job fit had a negative correlation with indicators of job strain. In fact, a primary intervention of career counseling can help to reduce stress levels (Firth-Cozens, 2003 ).
Job Redesign
The Job Demands-Control/Support (JD-C/S), Job Demands-Resources (JD-R), and transactional models all suggest that factors in the work context require modifications in order to reduce potential ill-health and poor organizational performance. Drawing on Hackman and Oldham’s ( 1980 ) Job Characteristics Model, it is possible to assess with the Job Diagnostics Survey (JDS) the current state of work characteristics related to skill variety, task identity, task significance, autonomy, and feedback. Modifying those aspects would help create a sense of meaningfulness, sense of responsibility, and feeling of knowing how one is performing, which subsequently affects a person’s well-being as identified in assessments of motivation, satisfaction, improved performance, and reduced withdrawal intentions and behaviors. Extending this argument to the stress models, it can be deduced that reducing uncertainty or perceived unfairness that may be associated with a person’s perception of these work characteristics, as well as making changes to physical characteristics of the environment (e.g., lighting, seating, desk, air quality), nature of work (e.g., job responsibilities, roles, decision-making latitude), and organizational arrangements (e.g., reporting structure and feedback mechanisms), can help mitigate against numerous ill-health consequences and reduced organizational performance. In fact, Fried et al. ( 2013 ) showed that healthy patients of a medical clinic whose jobs were excessively low (i.e., monotonous) or excessively high (i.e., overstimulating) on job enrichment (as measured by the JDS) had greater abdominal obesity than those whose jobs were optimally enriched. By taking stock of employees’ perceptions of the current work situation, managers might think about ways to enhance employees’ coping toolkit, such as training on how to deal with difficult clients or creating stimulating opportunities when jobs have low levels of enrichment.
Participatory Action Research Interventions
Participatory action research (PAR) is an intervention wherein, through group discussions, employees help to identify and define problems in organizational structure, processes, policies, practices, and reward structures, as well as help to design, implement, and evaluate success of solutions. PAR is in itself an intervention, but its goal is to design interventions to eliminate or reduce work-related factors that are impeding performance and causing people to be unwell. An example of a successful primary intervention, utilizing principles of PAR and driven by the JD-C and JD-C/S stress frameworks is Health Circles (HCs; Aust & Ducki, 2004 ).
HCs, developed in Germany in the 1980s, were popular practices in industries, such as metal, steel, and chemical, and service. Similar to other problem-solving practices, such as quality circles, HCs were based on the assumptions that employees are the experts of their jobs. For this reason, to promote employee well-being, management and administrators solicited suggestions and ideas from the employees to improve occupational health, thereby increasing employees’ job control. HCs also promoted communication between managers and employees, which had a potential to increase social support. With more control and support, employees would experience less strains and better occupational well-being.
Employing the three-steps of (1) problem analysis (i.e., diagnosis or discovery through data generated from organizational records of absenteeism length, frequency, rate, and reason and employee survey), (2) HC meetings (6 to 10 meetings held over several months to brainstorm ideas to improve occupational safety and health concerns identified in the discovery phase), and (3) HC evaluation (to determine if desired changes were accomplished and if employees’ reports of stressors and strains changed after the course of 15 months), improvements were to be expected (Aust & Ducki, 2004 ). Aust and Ducki ( 2004 ) reviewed 11 studies presenting 81 health circles in 30 different organizations. Overall study participants had high satisfaction with the HCs practices. Most companies acted upon employees’ suggestions (e.g., improving driver’s seat and cab, reducing ticket sale during drive, team restructuring and job rotation to facilitate communication, hiring more employees during summer time, and supervisor training program to improve leadership and communication skills) to improve work conditions. Thus, HCs represent a successful theory-grounded intervention to routinely improve employees’ occupational health.
Physical Setting
The physical environment or physical workspace has an enormous impact on individuals’ well-being, attitudes, and interactions with others, as well as on the implications on innovation and well-being (Oksanen & Ståhle, 2013 ; Vischer, 2007 ). In a study of 74 new product development teams (total of 437 study respondents) in Western Europe, Chong, van Eerde, Rutte, and Chai ( 2012 ) found that when teams were faced with challenge time pressures, meaning the teams had a strong interest and desire in tackling complex, but engaging tasks, when they were working proximally close with one another, team communication improved. Chong et al. assert that their finding aligns with prior studies that have shown that physical proximity promotes increased awareness of other team members, greater tendency to initiate conversations, and greater team identification. However, they also found that when faced with hindrance time pressures, physical proximity related to low levels of team communication, but when hindrance time pressure was low, team proximity had an increasingly greater positive relationship with team communication.
In addition to considering the type of work demand teams must address, other physical workspace considerations include whether people need to work collaboratively and synchronously or independently and remotely (or a combination thereof). Consideration needs to be given to how company contributors would satisfy client needs through various modes of communication, such as email vs. telephone, and whether individuals who work by a window might need shading to block bright sunlight from glaring on their computer screens. Finally, people who have to use the telephone for extensive periods of time would benefit from earphones to prevent neck strains. Most physical stressors are rather simple to rectify. However, companies are often not aware of a problem until after a problem arises, such as when a person’s back is strained from trying to move heavy equipment. Companies then implement strategies to remediate the environmental stressor. With the help of human factors, and organizational and office design consultants, many of the physical barriers to optimal performance can be prevented (Rousseau & Aubé, 2010 ). In a study of 215 French-speaking Canadian healthcare employees, Rousseau and Aubé ( 2010 ) found that although supervisor instrumental support positively related with affective commitment to the organization, the relationship was even stronger for those who reported satisfaction with the ambient environment (i.e., temperature, lighting, sound, ventilation, and cleanliness).
Secondary Interventions (Coping)
Secondary interventions, also referred to as coping, focus on resources people can use to mitigate the risk of work-related illness or workplace injury. Resources may include properties related to social resources, behaviors, and cognitive structures. Each of these resource domains may be employed to cope with stressors. Monat and Lazarus ( 1991 ) summarize the definition of coping as “an individual’s efforts to master demands (or conditions of harm, threat, or challenge) that are appraised (or perceived) as exceeding or taxing his or her resources” (p. 5). To master demands requires use of the aforementioned resources. Secondary interventions help employees become aware of the psychological, physical, and behavioral responses that may occur from the stressors presented in their working environment. Secondary interventions help a person detect and attend to stressors and identify resources for and ways of mitigating job strains. Often, coping strategies are learned skills that have a cognitive foundation and serve important functions in improving people’s management of stressors (Lazarus & Folkman, 1991 ). Coping is effortful, but with practice it becomes easier to employ. This idea is the foundation for understanding the role of resilience in coping with stressors. However, “not all adaptive processes are coping. Coping is a subset of adaptational activities that involves effort and does not include everything that we do in relating to the environment” (Lazarus & Folkman, 1991 , p. 198). Furthermore, sometimes to cope with a stressor, a person may call upon social support sources to help with tangible materials or emotional comfort. People call upon support resources because they help to restructure how a person approaches or thinks about the stressor.
Most secondary interventions are aimed at helping the individual, though companies, as a policy, might require all employees to partake in training aimed at increasing employees’ awareness of and skills aimed at handling difficult situations vis à vis company channels (e.g., reporting on sexual harassment or discrimination). Furthermore, organizations might institute mentoring programs or work groups to address various work-related matters. These programs employ awareness-raising activities, stress-education, or skills training (cf., Bhagat, Segovis, & Nelson, 2012 ), which include development of skills in problem-solving, understanding emotion-focused coping, identifying and using social support, and enhancing capacity for resilience. The aim of these programs, therefore, is to help employees proactively review their perceptions of psychological, physical, and behavioral job-related strains, thereby extending their resilience, enabling them to form a personal plan to control stressors and practice coping skills (Cooper, Dewe, & O’Driscoll, 2011 ).
Often these stress management programs are instituted after an organization has observed excessive absenteeism and work-related performance problems and, therefore, are sometimes categorized as a tertiary stress management intervention or even a primary (prevention) intervention. However, the skills developed for coping with stressors also place the programs in secondary stress management interventions. Example programs that are categorized as tertiary or primary stress management interventions may also be secondary stress management interventions (see Figure 1 ), and these include lifestyle advice and planning, stress inoculation training, simple relaxation techniques, meditation, basic trainings in time management, anger management, problem-solving skills, and cognitive-behavioral therapy. Corporate wellness programs also fall under this category. In other words, some programs could be categorized as primary, secondary, or tertiary interventions depending upon when the employee (or organization) identifies the need to implement the program. For example, time management practices could be implemented as a means of preventing some stressors, as a way to cope with mounting stressors, or as a strategy to mitigate symptoms of excessive of stressors. Furthermore, these programs can be administered at the individual level or group level. As related to secondary interventions, these programs provide participants with opportunities to develop and practice skills to cognitively reappraise the stressor(s); to modify their perspectives about stressors; to take time out to breathe, stretch, meditate, relax, and/or exercise in an attempt to support better decision-making; to articulate concerns and call upon support resources; and to know how to say “no” to onslaughts of requests to complete tasks. Participants also learn how to proactively identify coping resources and solve problems.
According to Cooper, Dewe, and O’Driscoll ( 2001 ), secondary interventions are successful in helping employees modify or strengthen their ability to cope with the experience of stressors with the goal of mitigating the potential harm the job stressors may create. Secondary interventions focus on individuals’ transactions with the work environment and emphasize the fit between a person and his or her environment. However, researchers have pointed out that the underlying assumption of secondary interventions is that the responsibility for coping with the stressors of the environment lies within individuals (Quillian-Wolever & Wolever, 2003 ). If companies cannot prevent the stressors in the first place, then they are, in part, responsible for helping individuals develop coping strategies and informing employees about programs that would help them better cope with job stressors so that they are able to fulfill work assignments.
Stress management interventions that help people learn to cope with stressors focus mainly on the goals of enabling problem-resolution or expressing one’s emotions in a healthy manner. These goals are referred to as problem-focused coping and emotion-focused coping (Folkman & Lazarus, 1980 ; Pearlin & Schooler, 1978 ), and the person experiencing the stressors as potential threat is the agent for change and the recipient of the benefits of successful coping (Hobfoll, 1998 ). In addition to problem-focused and emotion-focused coping approaches, social support and resilience may be coping resources. There are many other sources for coping than there is room to present here (see e.g., Cartwright & Cooper, 2005 ); however, the current literature has primarily focused on these resources.
Problem-Focused Coping
Problem-focused or direct coping helps employees remove or reduce stressors in order to reduce their strain experiences (Bhagat et al., 2012 ). In problem-focused coping employees are responsible for working out a strategic plan in order to remove job stressors, such as setting up a set of goals and engaging in behaviors to meet these goals. Problem-focused coping is viewed as an adaptive response, though it can also be maladaptive if it creates more problems down the road, such as procrastinating getting work done or feigning illness to take time off from work. Adaptive problem-focused coping negatively relates to long-term job strains (Higgins & Endler, 1995 ). Discussion on problem-solving coping is framed from an adaptive perspective.
Problem-focused coping is featured as an extension of control, because engaging in problem-focused coping strategies requires a series of acts to keep job stressors under control (Bhagat et al., 2012 ). In the stress literature, there are generally two ways to categorize control: internal versus external locus of control, and primary versus secondary control. Locus of control refers to the extent to which people believe they have control over their own life (Rotter, 1966 ). People high in internal locus of control believe that they can control their own fate whereas people high in external locus of control believe that outside factors determine their life experience (Rotter, 1966 ). Generally, those with an external locus of control are less inclined to engage in problem-focused coping (Strentz & Auerbach, 1988 ). Primary control is the belief that people can directly influence their environment (Alloy & Abramson, 1979 ), and thus they are more likely to engage in problem-focused coping. However, when it is not feasible to exercise primary control, people search for secondary control, with which people try to adapt themselves into the objective environment (Rothbaum, Weisz, & Snyder, 1982 ).
Emotion-Focused Coping
Emotion-focused coping, sometimes referred to as palliative coping, helps employees reduce strains without the removal of job stressors. It involves cognitive or emotional efforts, such as talking about the stressor or distracting oneself from the stressor, in order to lessen emotional distress resulting from job stressors (Bhagat et al., 2012 ). Emotion-focused coping aims to reappraise and modify the perceptions of a situation or seek emotional support from friends or family. These methods do not include efforts to change the work situation or to remove the job stressors (Lazarus & Folkman, 1991 ). People tend to adopt emotion-focused coping strategies when they believe that little or nothing can be done to remove the threatening, harmful, and challenging stressors (Bhagat et al., 2012 ), such as when they are the only individuals to have the skills to get a project done or they are given increased responsibilities because of the unexpected departure of a colleague. Emotion-focused coping strategies include (1) reappraisal of the stressful situation, (2) talking to friends and receiving reassurance from them, (3) focusing on one’s strength rather than weakness, (4) optimistic comparison—comparing one’s situation to others’ or one’s past situation, (5) selective ignoring—paying less attention to the unpleasant aspects of one’s job and being more focused on the positive aspects of the job, (6) restrictive expectations—restricting one’s expectations on job satisfaction but paying more attention to monetary rewards, (7) avoidance coping—not thinking about the problem, leaving the situation, distracting oneself, or using alcohol or drugs (e.g., Billings & Moos, 1981 ).
Some emotion-focused coping strategies are maladaptive. For example, avoidance coping may lead to increased level of job strains in the long run (e.g., Parasuraman & Cleek, 1984 ). Furthermore, a person’s ability to cope with the imbalance of performing work to meet organizational expectations can take a toll on the person’s health, leading to physiological consequences such as cardiovascular disease, sleep disorders, gastrointestinal disorders, and diabetes (Fried et al., 2013 ; Siegrist, 2010 ; Toker, Shirom, Melamed, & Armon, 2012 ; Willert, Thulstrup, Hertz, & Bonde, 2010 ).
Comparing Coping Strategies across Cultures
Most coping research is conducted in individualistic, Western cultures wherein emotional control is emphasized and both problem-solving focused coping and primary control are preferred (Bhagat et al., 2010 ). However, in collectivistic cultures, emotion-focused coping and use of secondary control may be preferred and may not necessarily carry a negative evaluation (Bhagat et al., 2010 ). For example, African Americans are more likely to use emotion-focused coping than non–African Americans (Knight, Silverstein, McCallum, & Fox, 2000 ), and among women who experienced sexual harassment, Anglo American women were less likely to employ emotion focused coping (i.e., avoidance coping) than Turkish women and Hispanic American women, while Hispanic women used more denial than the other two groups (Wasti & Cortina, 2002 ).
Thus, whereas problem-focused coping is venerated in Western societies, emotion-focused coping may be more effective in reducing strains in collectivistic cultures, such as China, Japan, and India (Bhagat et al., 2010 ; Narayanan, Menon, & Spector, 1999 ; Selmer, 2002 ). Indeed, Swedish participants reported more problem-focused coping than did Chinese participants (Xiao, Ottosson, & Carlsson, 2013 ), American college students engaged in more problem-focused coping behaviors than did their Japanese counterparts (Ogawa, 2009 ), and Indian (vs. Canadian) students reported more emotion-focused coping, such as seeking social support and positive reappraisal (Sinha, Willson, & Watson, 2000 ). Moreover, Glazer, Stetz, and Izso ( 2004 ) found that internal locus of control was more predominant in individualistic cultures (United Kingdom and United States), whereas external locus of control was more predominant in communal cultures (Italy and Hungary). Also, internal locus of control was associated with less job stress, but more so for nurses in the United Kingdom and United States than Italy and Hungary. Taken together, adoption of coping strategies and their effectiveness differ significantly across cultures. The extent to which a coping strategy is perceived favorably and thus selected or not selected is not only a function of culture, but also a person’s sociocultural beliefs toward the coping strategy (Morimoto, Shimada, & Ozaki, 2013 ).
Social Support
Social support refers to the aid an entity gives to a person. The source of the support can be a single person, such as a supervisor, coworker, subordinate, family member, friend, or stranger, or an organization as represented by upper-level management representing organizational practices. The type of support can be instrumental or emotional. Instrumental support, including informational support, refers to that which is tangible, such as data to help someone make a decision or colleagues’ sick days so one does not lose vital pay while recovering from illness. Emotional support, including esteem support, refers to the psychological boost given to a person who needs to express emotions and feel empathy from others or to have his or her perspective validated. Beehr and Glazer ( 2001 ) present an overview of the role of social support on the stressor-strain relationship and arguments regarding the role of culture in shaping the utility of different sources and types of support.
Meaningfulness and Resilience
Meaningfulness reflects the extent to which people believe their lives are significant, purposeful, goal-directed, and fulfilling (Glazer, Kożusznik, Meyers, & Ganai, 2014 ). When faced with stressors, people who have a strong sense of meaning in life will also try to make sense of the stressors. Maintaining a positive outlook on life stressors helps to manage emotions, which is helpful in reducing strains, particularly when some stressors cannot be problem-solved (Lazarus & Folkman, 1991 ). Lazarus and Folkman ( 1991 ) emphasize that being able to reframe threatening situations can be just as important in an adaptation as efforts to control the stressors. Having a sense of meaningfulness motivates people to behave in ways that help them overcome stressors. Thus, meaningfulness is often used in the same breath as resilience, because people who are resilient are often protecting that which is meaningful.
Resilience is a personality state that can be fortified and enhanced through varied experiences. People who perceive their lives are meaningful are more likely to find ways to face adversity and are therefore more prone to intensifying their resiliency. When people demonstrate resilience to cope with noxious stressors, their ability to be resilient against other stressors strengthens because through the experience, they develop more competencies (Glazer et al., 2014 ). Thus, fitting with Hobfoll’s ( 1989 , 2001 ) COR theory, meaningfulness and resilience are psychological resources people attempt to conserve and protect, and employ when necessary for making sense of or coping with stressors.
Tertiary Interventions (Stress Management)
Stress management refers to interventions employed to treat and repair harmful repercussions of stressors that were not coped with sufficiently. As Lazarus and Folkman ( 1991 ) noted, not all stressors “are amenable to mastery” (p. 205). Stressors that are unmanageable and lead to strains require interventions to reverse or slow down those effects. Workplace interventions might focus on the person, the organization, or both. Unfortunately, instead of looking at the whole system to include the person and the workplace, most companies focus on the person. Such a focus should not be a surprise given the results of van der Klink, Blonk, Schene, and van Dijk’s ( 2001 ) meta-analysis of 48 experimental studies conducted between 1977 and 1996 . They found that of four types of tertiary interventions, the effect size for cognitive-behavioral interventions and multimodal programs (e.g., the combination of assertive training and time management) was moderate and the effect size for relaxation techniques was small in reducing psychological complaints, but not turnover intention related to work stress. However, the effects of (the five studies that used) organization-focused interventions were not significant. Similarly, Richardson and Rothstein’s ( 2008 ) meta-analytic study, including 36 experimental studies with 55 interventions, showed a larger effect size for cognitive-behavioral interventions than relaxation, organizational, multimodal, or alternative. However, like with van der Klink et al. ( 2001 ), Richardson and Rothstein ( 2008 ) cautioned that there were few organizational intervention studies included and the impact of interventions were determined on the basis of psychological outcomes and not physiological or organizational outcomes. Van der Klink et al. ( 2001 ) further expressed concern that organizational interventions target the workplace and that changes in the individual may take longer to observe than individual interventions aimed directly at the individual.
The long-term benefits of individual focused interventions are not yet clear either. Per Giga, Cooper, and Faragher ( 2003 ), the benefits of person-directed stress management programs will be short-lived if organizational factors to reduce stressors are not addressed too. Indeed, LaMontagne, Keegel, Louie, Ostry, and Landsbergis ( 2007 ), in their meta-analysis of 90 studies on stress management interventions published between 1990 and 2005 , revealed that in relation to interventions targeting organizations only, and interventions targeting individuals only, interventions targeting both organizations and individuals (i.e. the systems approach) had the most favorable positive effects on both the organizations and the individuals. Furthermore, the organization-level interventions were effective at both the individual and organization levels, but the individual-level interventions were effective only at the individual level.
Individual-Focused Stress Management
Individual-focused interventions concentrate on improving conditions for the individual, though counseling programs emphasize that the worker is in charge of reducing “stress,” whereas role-focused interventions emphasize activities that organizations can guide to actually reduce unnecessary noxious environmental factors.
Individual-Focused Stress Management: Employee Assistance Programs
When stress become sufficiently problematic (which is individually gauged or attended to by supportive others) in a worker’s life, employees may utilize the short-term counseling services or referral services Employee Assistance Programs (EAPs) provide. People who utilize the counseling services may engage in cognitive behavioral therapy aimed at changing the way people think about the stressors (e.g., as challenge opportunity over threat) and manage strains. Example topics that may be covered in these therapy sessions include time management and goal setting (prioritization), career planning and development, cognitive restructuring and mindfulness, relaxation, and anger management. In a study of healthcare workers and teachers who participated in a 2-day to 2.5-day comprehensive stress management training program (including 26 topics on identifying, coping with, and managing stressors and strains), Siu, Cooper, and Phillips ( 2013 ) found psychological and physical improvements were self-reported among the healthcare workers (for which there was no control group). However, comparing an intervention group of teachers to a control group of teachers, the extent of change was not as visible, though teachers in the intervention group engaged in more mastery recovery experiences (i.e., they purposefully chose to engage in challenging activities after work).
Individual-Focused Stress Management: Mindfulness
A popular therapy today is to train people to be more mindful, which involves helping people live in the present, reduce negative judgement of current and past experiences, and practicing patience (Birnie, Speca, & Carlson, 2010 ). Mindfulness programs usually include training on relaxation exercises, gentle yoga, and awareness of the body’s senses. In one study offered through the continuing education program at a Canadian university, 104 study participants took part in an 8-week, 90 minute per group (15–20 participants per) session mindfulness program (Birnie et al., 2010 ). In addition to body scanning, they also listened to lectures on incorporating mindfulness into one’s daily life and received a take-home booklet and compact discs that guided participants through the exercises studied in person. Two weeks after completing the program, participants’ mindfulness attendance and general positive moods increased, while physical, psychological, and behavioral strains decreased. In another study on a sample of U.K. government employees, study participants receiving three sessions of 2.5 to 3 hours each training on mindfulness, with the first two sessions occurring in consecutive weeks and the third occurring about three months later, Flaxman and Bond ( 2010 ) found that compared to the control group, the intervention group showed a decrease in distress levels from Time 1 (baseline) to Time 2 (three months after first two training sessions) and Time 1 to Time 3 (after final training session). Moreover, of the mindfulness intervention study participants who were clinically distressed, 69% experienced clinical improvement in their psychological health.
Individual-Focused Stress Management: Biofeedback/Imagery/Meditation/Deep Breathing
Biofeedback uses electronic equipment to inform users about how their body is responding to tension. With guidance from a therapist, individuals then learn to change their physiological responses so that their pulse normalizes and muscles relax (Norris, Fahrion, & Oikawa, 2007 ). The therapist’s guidance might include reminders for imagery, meditation, body scan relaxation, and deep breathing. Saunders, Driskell, Johnston, and Salas’s ( 1996 ) meta-analysis of 37 studies found that imagery helped reduce state and performance anxiety. Once people have been trained to relax, reminder triggers may be sent through smartphone push notifications (Villani et al., 2013 ).
Smartphone technology can also be used to support weight loss programs, smoking cessation programs, and medication or disease (e.g., diabetes) management compliance (Heron & Smyth, 2010 ; Kannampallil, Waicekauskas, Morrow, Kopren, & Fu, 2013 ). For example, smartphones could remind a person to take medications or test blood sugar levels or send messages about healthy behaviors and positive affirmations.
Individual-Focused Stress Management: Sleep/Rest/Respite
Workers today sleep less per night than adults did nearly 30 years ago (Luckhaupt, Tak, & Calvert, 2010 ; National Sleep Foundation, 2005 , 2013 ). In order to combat problems, such as increased anxiety and cardiovascular artery disease, associated with sleep deprivation and insufficient rest, it is imperative that people disconnect from their work at least one day per week or preferably for several weeks so that they are able to restore psychological health (Etzion, Eden, & Lapidot, 1998 ; Ragsdale, Beehr, Grebner, & Han, 2011 ). When college students engaged in relaxation-type activities, such as reading or watching television, over the weekend, they experienced less emotional exhaustion and greater general well-being than students who engaged in resources-consuming activities, such as house cleaning (Ragsdale et al., 2011 ). Additional research and future directions for research are reviewed and identified in the work of Sonnentag ( 2012 ). For example, she asks whether lack of ability to detach from work is problematic for people who find their work meaningful. In other words, are negative health consequences only among those who do not take pleasure in their work? Sonnetag also asks how teleworkers detach from their work when engaging in work from the home. Ironically, one of the ways that companies are trying to help with the challenges of high workload or increased need to be available to colleagues, clients, or vendors around the globe is by offering flexible work arrangements, whereby employees who can work from home are given the opportunity to do so. Companies that require global interactions 24-hours per day often employ this strategy, but is the solution also a source of strain (Glazer, Kożusznik, & Shargo, 2012 )?
Individual-Focused Stress Management: Role Analysis
Role analysis or role clarification aims to redefine, expressly identify, and align employees’ roles and responsibilities with their work goals. Through role negotiation, involved parties begin to develop a new formal or informal contract about expectations and define resources needed to fulfill those expectations. Glazer has used this approach in organizational consulting and, with one memorable client engagement, found that not only were the individuals whose roles required deeper re-evaluation happier at work (six months later), but so were their subordinates. Subordinates who once characterized the two partners as hostile and akin to a couple going through a bad divorce, later referred to them as a blissful pair. Schaubroeck, Ganster, Sime, and Ditman ( 1993 ) also found in a three-wave study over a two-year period that university employees’ reports of role clarity and greater satisfaction with their supervisor increased after a role clarification exercise of top managers’ roles and subordinates’ roles. However, the intervention did not have any impact on reported physical symptoms, absenteeism, or psychological well-being. Role analysis is categorized under individual-focused stress management intervention because it is usually implemented after individuals or teams begin to demonstrate poor performance and because the intervention typically focuses on a few individuals rather than an entire organization or group. In other words, the intervention treats the person’s symptoms by redefining the role so as to eliminate the stimulant causing the problem.
Organization-Focused Stress Management
At the organizational level, companies that face major declines in productivity and profitability or increased costs related to healthcare and disability might be motivated to reassess organizational factors that might be impinging on employees’ health and well-being. After all, without healthy workers, it is not possible to have a healthy organization. Companies may choose to implement practices and policies that are expected to help not only the employees, but also the organization with reduced costs associated with employee ill-health, such as medical insurance, disability payments, and unused office space. Example practices and policies that may be implemented include flexible work arrangements to ensure that employees are not on the streets in the middle of the night for work that can be done from anywhere (such as the home), diversity programs to reduce stress-induced animosity and prejudice toward others, providing only healthy food choices in cafeterias, mandating that all employees have physicals in order to receive reduced prices for insurance, company-wide closures or mandatory paid time off, and changes in organizational visioning.
Organization-Focused Stress Management: Organizational-Level Occupational Health Interventions
As with job design interventions that are implemented to remediate work characteristics that were a source of unnecessary or excessive stressors, so are organizational-level occupational health (OLOH) interventions. As with many of the interventions, its placement as a primary or tertiary stress management intervention may seem arbitrary, but when considering the goal and target of change, it is clear that the intervention is implemented in response to some ailing organizational issues that need to be reversed or stopped, and because it brings in the entire organization’s workforce to address the problems, it has been placed in this category. There are several more case studies than empirical studies on the topic of whole system organizational change efforts (see example case studies presented by the United Kingdom’s Health and Safety Executive). It is possible that lack of published empirical work is not so much due to lack of attempting to gather and evaluate the data for publication, but rather because the OLOH interventions themselves never made it to the intervention stage, the interventions failed (Biron, Gatrell, & Cooper, 2010 ), or the level of evaluation was not rigorous enough to get into empirical peer-review journals. Fortunately, case studies provide some indication of the opportunities and problems associated with OLOH interventions.
One case study regarding Cardiff and Value University Health Board revealed that through focus group meetings with members of a steering group (including high-level managers and supported by top management) and facilitated by a neutral, non-judgemental organizational health consultant, ideas for change were posted on newsprint, discussed, and areas in the organization needing change were identified. The intervention for giving voice to people who initially had little already had a positive effect on the organization, as absence decreased by 2.09% and 6.9% merely 12 and 18 months, respectively, after the intervention. Translated in financial terms, the 6.9% change was equivalent to a quarterly savings of £80,000 (Health & Safety Executive, n.d. ). Thus, focusing on the context of change and how people will be involved in the change process probably helped the organization realize improvements (Biron et al., 2010 ). In a recent and rare empirical study, employing both qualitative and quantitative data collection methods, Sørensen and Holman ( 2014 ) utilized PAR in order to plan and implement an OLOH intervention over the course of 14 months. Their study aimed to examine the effectiveness of the PAR process in reducing workers’ work-related and social or interpersonal-related stressors that derive from the workplace and improving psychological, behavioral, and physiological well-being across six Danish organizations. Based on group dialogue, 30 proposals for change were proposed, all of which could be categorized as either interventions to focus on relational factors (e.g., management feedback improvement, engagement) or work processes (e.g., reduced interruptions, workload, reinforcing creativity). Of the interventions that were implemented, results showed improvements on manager relationship quality and reduced burnout, but no changes with respect to work processes (i.e., workload and work pace) perhaps because the employees already had sufficient task control and variety. These findings support Dewe and Kompier’s ( 2008 ) position that occupational health can be reinforced through organizational policies that reinforce quality jobs and work experiences.
Organization-Focused Stress Management: Flexible Work Arrangements
Dewe and Kompier ( 2008 ), citing the work of Isles ( 2005 ), noted that concern over losing one’s job is a reason for why 40% of survey respondents indicated they work more hours than formally required. In an attempt to create balance and perceived fairness in one’s compensation for putting in extra work hours, employees will sometimes be legitimately or illegitimately absent. As companies become increasingly global, many people with desk jobs are finding themselves communicating with colleagues who are halfway around the globe and at all hours of the day or night (Glazer et al., 2012 ). To help minimize the strains associated with these stressors, companies might devise flexible work arrangements (FWA), though the type of FWA needs to be tailored to the cultural environment (Masuda et al., 2012 ). FWAs give employees some leverage to decide what would be the optimal work arrangement for them (e.g., part-time, flexible work hours, compressed work week, telecommuting). In other words, FWA provides employees with the choice of when to work, where to work (on-site or off-site), and how many hours to work in a day, week, or pay period (Kossek, Thompson, & Lautsch, 2015 ). However, not all employees of an organization have equal access to or equitable use of FWAs; workers in low-wage, hourly jobs are often beholden to being physically present during specific hours (Swanberg McKechnie, Ojha, & James, 2011 ). In a study of over 1,300 full-time hourly retail employees in the United States, Swanberg et al. ( 2011 ) showed that employees who have control over their work schedules and over their work hours were satisfied with their work schedules, perceived support from the supervisor, and work engagement.
Unfortunately, not all FWAs yield successful results for the individual or the organization. Being able to work from home or part-time can have problems too, as a person finds himself or herself working more hours from home than required. Sometimes telecommuting creates work-family conflict too as a person struggles to balance work and family obligations while working from home. Other drawbacks include reduced face-to-face contact between work colleagues and stakeholders, challenges shaping one’s career growth due to limited contact, perceived inequity if some have more flexibility than others, and ambiguity about work role processes for interacting with employees utilizing the FWA (Kossek et al., 2015 ). Organizations that institute FWAs must carefully weigh the benefits and drawbacks the flexibility may have on the employees using it or the employees affected by others using it, as well as the implications on the organization, including the vendors who are serving and clients served by the organization.
Organization-Focused Stress Management: Diversity Programs
Employees in the workplace might experience strain due to feelings of discrimination or prejudice. Organizational climates that do not promote diversity (in terms of age, religion, physical abilities, ethnicity, nationality, sex, and other characteristics) are breeding grounds for undesirable attitudes toward the workplace, lower performance, and greater turnover intention (Bergman, Palmieri, Drasgow, & Ormerod, 2012 ; Velez, Moradi, & Brewster, 2013 ). Management is thus advised to implement programs that reinforce the value and importance of diversity, as well as manage diversity to reduce conflict and feelings of prejudice. In fact, managers who attended a leadership training program reported higher multicultural competence in dealing with stressful situations (Chrobot-Mason & Leslie, 2012 ), and managers who persevered through challenges were more dedicated to coping with difficult diversity issues (Cilliers, 2011 ). Thus, diversity programs can help to reduce strains by directly reducing stressors associated with conflict linked to diversity in the workplace and by building managers’ resilience.
Organization-Focused Stress Management: Healthcare Management Policies
Over the past few years, organizations have adopted insurance plans that implement wellness programs for the sake of managing the increasing cost of healthcare that is believed to be a result of individuals’ not managing their own health, with regular check-ups and treatment. The wellness programs require all insured employees to visit a primary care provider, complete a health risk assessment, and engage in disease management activities as specified by a physician (e.g., see frequently asked questions regarding the State of Maryland’s Wellness Program). Companies believe that requiring compliance will reduce health problems, although there is no proof that such programs save money or that people would comply. One study that does, however, boast success, was a 12-week workplace health promotion program aimed at reducing Houston airport workers’ weight (Ebunlomo, Hare-Everline, Weber, & Rich, 2015 ). The program, which included 235 volunteer participants, was deemed a success, as there was a total weight loss of 345 pounds (or 1.5 lbs per person). Given such results in Houston, it is clear why some people are also skeptical over the likely success of wellness programs, particularly as there is no clear method for evaluating their efficacy (Sinnott & Vatz, 2015 ).
Moreover, for some, such a program is too paternalistic and intrusive, as well as punishes anyone who chooses not to actively participate in disease management programs (Sinnott & Vatz, 2015 ). The programs put the onus of change on the person, though it is a response to the high costs of ill-health. The programs neglect to consider the role of the organization in reducing the barriers to healthy lifestyle, such as cloaking exempt employment as simply needing to get the work done, when it usually means working significantly more hours than a standard workweek. In fact, workplace health promotion programs did not reduce presenteeism (i.e., people going to work while unwell thereby reducing their job performance) among those who suffered from physical pain (Cancelliere, Cassidy, Ammendolia, & Côte, 2011 ). However, supervisor education, worksite exercise, lifestyle intervention through email, midday respite from repetitive work, a global stress management program, changes in lighting, and telephone interventions helped to reduce presenteeism. Thus, emphasis needs to be placed on psychosocial aspects of the organization’s structure, including managers and overall organizational climate for on-site presence, that reinforces such behavior (Cancelliere et al., 2011 ). Moreover, wellness programs are only as good as the interventions to reduce work-related stressors and improve organizational resources to enable workers to improve their overall psychological and physical health.
Concluding Remarks
Future research.
One of the areas requiring more theoretical and practical attention is that of the utility of stress frameworks to guide organizational development change interventions. Although it has been proposed that the foundation for work stress management interventions is in organizational development, and even though scholars and practitioners of organization development were also founders of research programs that focused on employee health and well-being or work stress, there are few studies or other theoretical works that link the two bodies of literature.
A second area that requires additional attention is the efficacy of stress management interventions across cultures. In examining secondary stress management interventions (i.e., coping), some cross-cultural differences in findings were described; however, there is still a dearth of literature from different countries on the utility of different prevention, coping, and stress management strategies.
A third area that has been blossoming since the start of the 21st century is the topic of hindrance and challenge stressors and the implications of both on workers’ well-being and performance. More research is needed on this topic in several areas. First, there is little consistency by which researchers label a stressor as a hindrance or a challenge. Researchers sometimes take liberties with labels, but it is not the researchers who should label a stressor but the study participants themselves who should indicate if a stressor is a source of strain. Rodríguez, Kozusznik, and Peiró ( 2013 ) developed a measure in which respondents indicate whether a stressor is a challenge or a hindrance. Just as some people may perceive demands to be challenges that they savor and that result in a psychological state of eustress (Nelson & Simmons, 2003 ), others find them to be constraints that impede goal fulfillment and thus might experience distress. Likewise, some people might perceive ambiguity as a challenge that can be overcome and others as a constraint over which he or she has little control and few or no resources with which to cope. More research on validating the measurement of challenge vs. hindrance stressors, as well as eustress vs. distress, and savoring vs. coping, is warranted. Second, at what point are challenge stressors harmful? Just because people experiencing challenge stressors continue to perform well, it does not necessarily mean that they are healthy people. A great deal of stressors are intellectually stimulating, but excessive stimulation can also take a toll on one’s physiological well-being, as evident by the droves of professionals experiencing different kinds of diseases not experienced as much a few decades ago, such as obesity (Fried et al., 2013 ). Third, which stress management interventions would better serve to reduce hindrance stressors or to reduce strain that may result from challenge stressors while reinforcing engagement-producing challenge stressors?
A fourth area that requires additional attention is that of the flexible work arrangements (FWAs). One of the reasons companies have been willing to permit employees to work from home is not so much out of concern for the employee, but out of the company’s need for the focal person to be able to communicate with a colleague working from a geographic region when it is night or early morning for the focal person. Glazer, Kożusznik, and Shargo ( 2012 ) presented several areas for future research on this topic, noting that by participating on global virtual teams, workers face additional stressors, even while given flexibility of workplace and work time. As noted earlier, more research needs to be done on the extent to which people who take advantage of FWAs are advantaged in terms of detachment from work. Can people working from home detach? Are those who find their work invigorating also likely to experience ill-health by not detaching from work?
A fifth area worthy of further research attention is workplace wellness programing. According to Page and Vella-Brodrick ( 2009 ), “subjective and psychological well-being [are] key criteria for employee mental health” (p. 442), whereby mental health focuses on wellness, rather than the absence of illness. They assert that by fostering employee mental health, organizations are supporting performance and retention. Employee well-being can be supported by ensuring that jobs are interesting and meaningful, goals are achievable, employees have control over their work, and skills are used to support organizational and individual goals (Dewe & Kompier, 2008 ). However, just as mental health is not the absence of illness, work stress is not indicative of an absence of psychological well-being. Given the perspective that employee well-being is a state of mind (Page & Vella-Brodrick, 2009 ), we suggest that employee well-being can be negatively affected by noxious job stressors that cannot be remediated, but when job stressors are preventable, employee well-being can serve to protect an employee who faces job stressors. Thus, wellness programs ought to focus on providing positive experiences by enhancing and promoting health, as well as building individual resources. These programs are termed “green cape” interventions (Pawelski, 2016 ). For example, with the growing interests in positive psychology, researchers and practitioners have suggested employing several positive psychology interventions, such as expressing gratitude, savoring experiences, and identifying one’s strengths (Tetrick & Winslow, 2015 ). Another stream of positive psychology is psychological capital, which includes four malleable functions of self-efficacy, optimism, hope, and resilience (Luthans, Youssef, & Avolio, 2007 ). Workplace interventions should include both “red cape” interventions (i.e., interventions to reduce negative experiences) and “green cape” interventions (i.e., workplace wellness programs; Polly, 2014 ).
A Healthy Organization’s Pledge
A healthy workplace requires healthy workers. Period. Among all organizations’ missions should be the focus on a healthy workforce. To maintain a healthy workforce, the company must routinely examine its own contributions in terms of how it structures itself; reinforces communications among employees, vendors, and clients; how it rewards and cares for its people (e.g., ensuring they get sufficient rest and can detach from work); and the extent to which people at the upper levels are truly connected with the people at the lower levels. As a matter of practice, management must recognize when employees are overworked, unwell, and poorly engaged. Management must also take stock of when it is doing well and right by its contributors’ and maintain and reinforce the good practices, norms, and procedures. People in the workplace make the rules; people in the workplace can change the rules. How management sees its employees and values their contribution will have a huge role in how a company takes stock of its own pain points. Providing employees with tools to manage their own reactions to work-related stressors and consequent strains is fine, but wouldn’t it be grand if organizations took better notice about what they could do to mitigate the strain-producing stressors in the first place and take ownership over how employees are treated?
- Abramis, D. J. (1994). Work role ambiguity, job satisfaction, and job performance: Meta-analyses and review. Psychological Report, 75 , 1411–1433.
- Adams, G. A. , & Jex, S. M. (1999). Relationships between time management, control, work–family conflict, and strain. Journal of Occupational Health Psychology, 1 , 72–77.
- Alloy, L. B. , & Abramson, L. Y. (1979). Judgment of contingency in depressed and nondepressed students: Sadder but wiser? Journal of Experimental Psychology: General, 108 , 441–483.
- Amstad, F. T. , Meier, L. L. , Fasel, U. , Elfering, A. , & Semmer, N. K. (2011). A meta-analysis of work-family conflict and various outcomes with a special emphasis on cross-domain versus matching-domain relations. Journal of Occupational Health Psychology, 16 , 151–169.
- Assouline, M. , & Meir, E. I. (1987). Meta-analysis of the relationship between congruence and well-being measures. Journal of Vocational Behavior, 31 , 319–332.
- Aust, B. , & Ducki, A. (2004). Comprehensive health promotion interventions at the workplace: Experiences with health circles in Germany. Journal of Occupational Health Psychology, 9 , 258–270.
- Bakker, A. B. Demerouti, E. , Sanz-Vergel, A. I. (2014). Burnout and work engagement: The JD-R approach. Annual Review of Organizational Behavior, 1 , 389–411.
- Bakker, A. B. , van Veldhoven, M. J. P. M. , & Xanthopoulou, D. (2010). Beyond the demand-control model: Thriving on high job demands and resources. Journal of Personnel Psychology, 9 , 3–16.
- Beehr, T. A. , Glaser, K. M. , Canali, K. G. , & Wallwey, D. A. (2001). Back to basics: Re-examination of demand control theory of occupational stress. Work & Stress, 15 , 115–130.
- Beehr, T. A. , & Glazer, S. (2001). A cultural perspective of social support in relation to occupational stress. In P. Perrewé , D. C. Ganster , & J. Moran (Eds.), Research in occupational stress and well-being (pp. 97–142). Amsterdam: JAI Press.
- Bergman, M. E. , Palmieri, P. A. , Drasgow, F. , & Ormerod, A. J. (2012). Racial/ethnic harassment and discrimination, its antecedents, and its effect on job-related outcomes. Journal of Occupational Health Psychology, 17 , 65–78.
- Bhagat, R. S. , Krishnan, B. , Nelson, T. A. , Leonard, K. M. , Ford, D. J. , & Billing, T. K. (2010). Organizational stress, psychological strain, and work outcomes in six national contexts: A closer look at the moderating influences of coping styles and decision latitude . Cross Cultural Management, 17 , 10–29.
- Bhagat, R. S. , O’Driscoll, M. P. , Babakus, E. , Frey, L. , Chokkar, J. , Ninokumar, B. H , et al. (1994). Organizational stress and coping in seven national contexts: A cross-cultural investigation . In G. P. Keita & J. J. Hurrell (Eds.), Job stress in a changing workforce: Investigating gender, diversity, and family issues (pp. 93–105). Washington, DC: American Psychological Association.
- Bhagat, R. S. , Segovis, J. C. , & Nelson, T. A. (2012). Work stress and coping in the era of globalization . New York: Routledge.
- Billings, A. G. , & Moos, R. H. (1981). The role of coping responses and social resources in attenuating the stress of life events. Journal of Behavioral Medicine, 4 , 139–157.
- Birnie, K. , Speca, M. , & Carlson, L. E. (2010). Exploring self-compassion and empathy in the context of mindfulness-based stress reduction (MBSR). Stress & Health, 26 , 359–371.
- Biron, C. , Gatrell, C. , & Cooper, C. L. (2010). Autopsy of a failure: Evaluating process and contextual issues in an organizational-level work stress intervention. International Journal of Stress Management, 17 , 135–158.
- Bowling, N. A. , & Kirkendall, C. (2012). Workload: A review of causes, consequences, and potential interventions. In J. Houdmont , S. Leka , & R. Sinclair (Eds.), Contemporary occupational health psychology (Vol. 2) (pp. 221–238). Chichester, U.K.: Wiley.
- Breaugh, J. A. , & Colihan, J. P. (1994). Measuring facets of job ambiguity: Construct validity evidence. Journal of Applied Psychology, 79 , 191–202.
- Bruk-Lee, V. , & Spector, P. E. (2006). The social stressors-counterproductive work behaviors link: Are conflicts with supervisors and coworkers the same? . Journal of Occupational Health Psychology, 11 , 145–156.
- Cancelliere, C. , Cassidy, J. D. , Ammendolia, C. , & Côte, P. (2011). Are workplace health promotion programs effective at improving presenteeism in workers? A systematic review and best evidence synthesis of the literature. BMC Public Health, 11 , 395–406.
- Caplan, R. D. (1987). Person–environment fit in organizations: Theories, facts, and values. In A. W. Riley & S. J. Zaccaro (Eds.), Occupational stress and organizational effectiveness (pp. 103–140). New York: Praeger.
- Cartwright, S. , & Cooper, C. L. (2005). Individually targeted interventions. In J. Barling , E. K. Kelloway , & M. R. Frone (Eds.), Handbook of work stress (pp. 607–622). Thousand Oaks, CA: SAGE.
- Cavanaugh, M. A. , Boswell, W. R. , Roehling, M. V. , & Boudreau, J. W. (2000). An empirical examination of self-reported work stress among U.S. managers . Journal of Applied Psychology, 85 , 65–74.
- Chong, D. S. F. , van Eerde, W. , Rutte, C. G. , & Chai, K. H. (2012). Bringing employees closer: The effect of proximity on communication when teams function under time pressure. Journal of Product Innovation Management, 29 , 205–215.
- Chrobot-Mason, D. , & Leslie, J. B. (2012). The role of multicultural competence and emotional intelligence in managing diversity. Psychologist-Manager Journal, 15 , 219–236.
- Cilliers, F. (2011). Individual diversity management and salutogenic functioning. International Review of Psychiatry, 23 , 501–507.
- Claessens, B. C. , Van Eerde, W. , Rutte, C. G. , & Roe, R. A. (2004). Planning behavior and perceived control of time at work . Journal of Organizational Behavior, 25 , 937–950.
- Cooper, C. L. , Dewe, P. D. , & O’Driscoll, M. P. (2011). Employee assistance programs: Strengths, challenges, and future roles. In J. C. Quick , L. E. Tetrick , J. C. Quick , L. E. Tetrick (Eds.), Handbook of occupational health psychology (2d ed.) (pp. 337–356). Washington, DC: American Psychological Association.
- Cooper, C. L. , Dewe, P. J. , & O’Driscoll, M. P. (2001). Organizational stress: A review and critique of theory, research, and applications . Thousand Oaks, CA: SAGE.
- Coovert, M. D. & Thompson, L. F. (2003). Technology and workplace health. In J. C. Quick & L. E. Tetrick (Eds.), Handbook of occupational health psychology (pp. 221–241). Washington, DC: American Psychological Association.
- Dawson, K. M. , O’Brien, K. E. , & Beehr, T. A. (2016). The role of hindrance stressors in the job demand-control-support model of occupational stress: A proposed theory revision . Journal of Organizational Behavior, 37 (3), 397–415.
- Dewe, P. , & Kompier, M. (2008). Foresight mental capital and wellbeing project: Wellbeing and work: Future challenges . London: The Government Office for Science.
- Dopkeen, J. C. , & DuBois, R. (2014). Stress in the workplace: A policy synthesis on its dimensions and prevalence . White paper. University of Illinois Chicago, Center for Employee Health Studies, School of Public Health.
- Dwyer, D. J. , & Ganster, D. C. (1991). The effects of job demands and control on employee attendance and satisfaction. Journal of Organizational Behavior, 12 , 595–608.
- Ebunlomo, E. O. , Hare-Everline, N. , Weber, A. , & Rich, J. (2015). Development of a comprehensive 12-week health promotion program for Houston Airport System. Texas Public Health Journal, 67 (1), 11–13.
- Edwards, J. R. (2008). Person-environment fit in organizations: An assessment of theoretical progress. The Academy of Management Annals, 2 , 167–230.
- Etzion, D. , Eden, D. , & Lapidot, Y. (1998). Relief from job stressors and burnout: Reserve service as a respite. Journal of Applied Psychology, 83 , 577–585.
- Firth-Cozens, J. (2003). Doctors, their wellbeing, and their stress: It’s time to be proactive about stress—and prevent it. British Medical Journal, 326 , 670–671.
- Flaxman, P. E. , & Bond, F. W. (2010). Worksite stress management training: Moderated effects and clinical significance. Journal of Occupational Health Psychology, 15 , 347–358.
- Folkman, S. , & Lazarus, R. S. (1980). An analysis of coping in a middle-aged community sample . Journal of Health and Social Behavior, 21 , 219–239.
- French, J. R. P., Jr. , & Caplan, R. D. (1972). Organizational stress and individual strain. In A. Marrow (Ed.), The failure of success . New York: AMACOM.
- French, J. R. P., Jr. , & Kahn, R. L. (1962). A programmatic approach to studying the industrial environment and mental health. Journal of Social Issues, 18 , 1–48.
- French, S. (2015, May 27). PCS workload and work-life balance survey 2013 . London: Public and Commercial Services Union.
- Fried, Y. , Laurence, G. A. , Shirom, A. , Melamed, S. , Toker, S. , Berliner, S. , & Shapira, I. (2013). The relationship between job enrichment and abdominal obesity: A longitudinal field study of apparently healthy individuals. Journal of Occupational Health Psychology, 18 , 458–468.
- Frone, M. R. (2000). Interpersonal conflict at work and psychological outcomes: Testing a model among young workers. Journal of Occupational Health Psychology, 5 , 246–255.
- Giga, S. I. , Cooper, C. L. , & Faragher, B. (2003). The development of a framework for a comprehensive approach to stress management interventions at work. International Journal of Stress Management, 10 , 280–296.
- Glazer, S. (2008). Cross-cultural issues in stress and burnout. In J. R. B. Halbesleben (Ed.), Handbook of Stress and Burnout in Health Care (pp. 79–93). Huntington, NY: Nova Science Publishers.
- Glazer, S. , & Beehr, T. A. (2005). Consistency of the implications of three role stressors across four countries. Journal of Organizational Behavior , 26 , 467–487.
- Glazer, S. , Kożusznik, M. W. , Meyers, J. H. , & Ganai, O. (2014). Cultural implications of meaningfulness as a resource to mitigate work stress. In S. Leka & R. Sinclair (Eds.), Contemporary occupational health psychology: Global perspectives on research and practice (Vol. 3) (pp. 114–130). Hoboken, NJ: Wiley.
- Glazer, S. , Kożusznik, M. W. , & Shargo, I. A. (2012). Global virtual teams: A cure for- or a cause of- stress. In P. L. Perrewé , J. Halbesleben , & C. Rosen (Eds.), Research in occupational stress and well being: The role of the economic context on occupational stress and well being (Vol. 10, pp. 213–266). Bingley, U.K.: Emerald.
- Glazer, S. , Stetz, T. A. , & Izso, L. (2004). Effects of personality on subjective job stress: A cultural analysis. Personality and Individual Differences, 37 , 645–658.
- Greenhaus, J. H. , & Beutell, N. J. (1985). Sources of conflict between work and family roles. Academy of Management Review, 10 , 76–88.
- Hackman, J. R. , & Oldham, G. R. (1980). Work Redesign . Reading, MA: Addison-Wesley.
- Hauke, A. , Flintrop, J. , Brun, E. , & Rugulies, R. (2011). The impact of work-related psychosocial stressors on the onset of musculoskeletal disorders in specific body regions: A review and meta-analysis of 54 longitudinal studies . Work & Stress, 25 , 243–256.
- Health & Safety Executive (n.d.). Cardiff and Value University Health Board—A stress case study .
- Heron, K. E. , & Smyth, J. M. (2010). Ecological momentary interventions: Incorporating mobile technology into psychosocial and health behaviour treatments. British Journal of Health Psychology, 15 , 1–39.
- Hershcovis, M. (2011). “Incivility, social undermining, bullying … oh my!”: A call to reconcile constructs within workplace aggression research . Journal of Organizational Behavior, 32 , 499–519.
- Higgins, J. E. , & Endler, N. S. (1995). Coping, life stress, and psychological and somatic distress . European Journal of Personality, 9 , 253–270.
- Hobfoll, S. E. (1989). Conservation of resources: A new attempt at conceptualizing stress. American Psychologist, 44 , 513–524.
- Hobfoll, S. E. (1998). Stress, culture, and community: The psychology and philosophy of stress . New York: Plenum Press.
- Hobfoll, S. E. (2001). The influence of culture, community, and the nested-self in the stress process: Advancing Conservation of Resources Theory. Applied Psychology: An International Review, 50 , 337–421.
- International Labor Organization (2012, July 5). Why stress at work matters . International Labor Organization.
- International Labor Organization (n.d.). Psychosocial risks and work-related stress . International Labor Organization.
- Jackson, S. E. , & Schuler, R. S. (1985). A meta-analysis and conceptual critique of research on role ambiguity and role conflict in work settings. Organizational Behavior and Human Decision Processes, 36 , 16–78.
- Jayson, S. (2012, January 11). Yeah, we’re stressed but dealing with it; Americans report a decrease in stress for the first time in five years, maybe because it’s just the new normal . USA Today .
- Jex, S. M. , & Beehr, T. A. (1991). Emerging theoretical and methodological issues in the study of work-related stress. Research in Personnel and Human Resources Management, 9 , 311–365.
- Jex, S. M. , & Elaqua, T. C. (1999). Time management as a moderator of relations between stressors and employee strain. Work & Stress, 13 , 182–191.
- Johnson, J. V. , & Hall, E. M. (1988). Job strain, workplace social support, and cardiovascular disease: A cross-sectional study of a random sample of the Swedish working population. American Journal of Public Health, 78 , 1336–1342.
- Kahn, R. L. , Wolfe, D. M. , Quinn, R. P. , Snoek, J. D. , & Rosenthal, R. A. (1964). Organizational stress: Studies in role conflict and ambiguity . New York: Wiley.
- Kannampallil, T. G. , Waicekauskas, K. , Morrow, D. G. , Kopren, K. M. , & Fu, W. (2013). External tools for collaborative medication scheduling. Cognition, Technology & Work, 15 , 121–131.
- Karasek, R. A. (1979). Job demands, job decision latitude, and mental strain: Implications for job redesign. Administrative Science Quarterly , 24 , 285–308.
- Karasek, R. A. , & Theorell, T. (1990). Healthy work: Stress, productivity, and the reconstruction of working life . New York: Basic Books.
- van der Klink, J. J. L. , Blonk, R. W. B. , Schene, A. H. , & van Dijk, F. J. H. (2001). The benefits of interventions for work-related stress. American Journal of Public Health, 91 , 270–276.
- Knight, B. G. , Silverstein, M. , McCallum, T. J. , & Fox, L. S. (2000). A sociocultural stress and coping model for mental health outcomes among African American caregivers in southern California . The Journals of Gerontology: Series B: Psychological Sciences and Social Sciences, 55B , 142–150.
- Kossek, E. E. , Thompson, R. J. , Lautsch, B. A. (2015). Balanced workplace flexibility: Avoiding the traps. California Management Review, 57 , 5–25.
- Kożusznik, M. , Rodriguez, I. , & Peiró, J. M. (2012). Cross-national outcomes of stress appraisal. Cross Cultural Management, 19 , 507–525.
- Kożusznik, M. , Rodriguez, I. , & Peiró, J. M. (2015). Eustress and distress climates in teams: Patterns and outcomes. International Journal of Stress Management, 22 , 1–23.
- Kristof-Brown, A. L. , Zimmerman, R. D. , & Johnson, E. C. (2005). Consequences of individuals’ fit at work: A meta-analysis of person-job, person-organization, person-group, and person-supervisor fit. Personnel Psychology, 58 , 281–342.
- LaMontagne, A. D. , Keegel, T. , Louie, A. M. , Ostry, A. , & Landsbergis, P. A. (2007). A systematic review of the job-stress intervention evaluation literature, 1990–2005. International Journal of Occupational and Environmental Health, 13 , 268–280.
- LaRocco, J. M. , Tetrick, L. E. , & Meder, D. (1989). Differences in perceptions of work environment conditions, job attitudes, and health beliefs among military physicians, dentists, and nurses. Military Psychology, 1 , 135–151.
- Lazarus, R. S. , & Folkman, S. (1984). Stress, appraisal, and coping . New York: Springer.
- Lazarus, R. S. , & Folkman, S. (1991). The concept of coping. In A. Monat & R. S. Lazarus (Eds.), Stress and coping: An anthology (3d ed.) (pp. 189–206). New York: Columbia University Press.
- LePine, J. A. , Podsakoff, N. P. , & LePine, M. A. (2005). A meta-analytic test of the challenge stressor-hindrance stressor framework: An explanation for inconsistent relationships among stressors and performance. Academy of Management Journal, 48 , 764–775.
- Lewin, K. (1936). Principles of topological psychology . New York: McGraw-Hill.
- Lewin K. (1951). Field theory in social science . New York: Harper and Row.
- Lewin, K. (1997). Resolving social conflicts & Field theory in social science . Washington, DC: American Psychological Association. Previously published in 1948 and 1951.
- Lin, B. C. , Kain, J. M. , & Fritz, C. (2013). Don’t interrupt me! An examination of the relationship between intrusions at work and employee strain. International Journal of Stress Management, 20 , 77–94.
- Liu, C. , & Li, H. (2017) Stressor and stressor appraisals: The moderating effect of task efficacy . Journal of Business and Psychology , 1–14.
- Liu, C. , Liu, Y. , Spector, P. E. , Shi, L. (2011). The interaction of job autonomy and conflict with supervisor in China and the United States: A qualitative and quantitative comparison. International Journal of Stress Management, 18 , 222–245.
- Liu, C. , Spector, P. E. , & Shi, L. (2007). Cross-national job stress: A quantitative and qualitative study. Journal of Organizational Behavior, 28 , 209–239.
- Luckhaupt, S. E. , Tak, S. , Calvert, G. M. (2010). The prevalence of short sleep duration by industry and occupational in the National Health Interview Survey. Sleep, 33 , 149–159.
- Luthans, F. , Youssef, C. M. , & Avolio, B. J. (2007). Psychological capital: Developing the human competitive edge . New York: Oxford University Press.
- Macan, T. H. (1994). Time management: Test of a process model. Journal of Applied Psychology, 79 , 381–391.
- Macan, T. H. , Shahani, C. , Dipboye, R. L. , & Philips, A. P. (1990). College students’ time management: Correlations with academic performance and stress. Journal of Educational Psychology, 82 , 760–768.
- Masuda, A. D. , Poelmans, S. A. Y. , Allen, T. D. , Spector, P. E. , Lapierre, L. M. , Cooper, C. L. , et al. (2012). Flexible work arrangements availability and their relationship with work-to-family conflict, job satisfaction, and turnover intentions: A comparison of three country clusters. Applied Psychology: An International Review, 61 , 1–29.
- McMahon, M. (2007). Think you might be addicted to email? You’re not alone . AOL.
- Monat, A. , & Lazarus, R. S. (Eds.). (1991). Stress and coping: An anthology (3d ed.). New York: Columbia University Press.
- Morimoto, H. , Shimada, H. , & Ozaki, K. (2013). Does stressor evaluation mediate sociocultural influence on coping selection? An investigation using Japanese employees. International Journal of Stress Management, 20 , 1–19.
- Narayanan, L. , Menon, S. , & Spector, P. E. (1999). A cross-cultural comparison of job stressors and reactions among employees holding comparable jobs in two countries. International Journal of Stress Management, 6 , 197–212.
- National Sleep Foundation (2005). Segment profiles . National Sleep Foundation.
- National Sleep Foundation (2013). How much sleep do adults need? . National Sleep Foundation.
- Nelson, D. L. , & Simmons, B. L. (2003). Health psychology and work stress: A more positive approach. In J. C. Quick & L. E. Tetrick (Eds.), Handbook of Occupational Health Psychology (pp. 97–119). Washington, DC: American Psychological Association.
- Norris, P. A. , Fahrion, S. L. , & Oikawa, L. O. (2007). Autogenic biofeedback training in psychophysiological therapy and stress management. In P. M. Lehrer , R. L. Woolfolk , & W. E. Sime (Eds.), Principles and practices of stress management (3d ed., pp. 175–205). New York: Guilford.
- Ogawa, N. (2009). Stress, coping behavior, and social support in Japan and the United States. Dissertation Abstracts International Section A , 69 , 3802.
- Oksanen, K. , & Ståhle, P. (2013). Physical environment as a source for innovation: Investigating the attributes of innovative space. Journal of Knowledge Management, 17 , 815–827.
- Page, K. M. , & Vella-Brodrick, D. A. (2009). The “what,” “why” and “how” of employee well-being: A new model. Social Indicators Research, 90 , 441–458.
- Parasuraman, S. , & Cleek, M. A. (1984). Coping behaviors and managers’ affective reactions to role stressors . Journal of Vocational Behavior, 24 , 179–193.
- Parasuraman, S. , & Purohit, Y. S. (2000). Distress and boredom among orchestra musicians: The two faces of stress. Journal of Occupational Health Psychology, 5 , 74–83.
- Pawelski, J. O. (2016). Defining the “positive” in positive psychology: Part II. A normative analysis . Journal of Positive Psychology, 11 , 357–365.
- Pearlin, L. I. , & Schooler, C. (1978). The structure of coping . Journal of Health and Social Behavior, 19 , 2–21.
- Peeters, M. A. G. , & Rutte, C. G. (2005). Time management behavior as a moderator for the job demand-control interaction. Journal of Occupational Health Psychology , 1 , 64–75.
- Peters, L. H. , & O’Connor, E. J. (1980). Situational constraints and work outcomes: The influence of a frequently overlooked construct. Academy of Management Review , 5, 391–397.
- Peters, L. H. , & O’Connor, E. J. (1988). Measuring work obstacles: Procedures, issues, and implications. In F. D. Schoorman & B. Schneider (Eds.), Facilitating work effectiveness (pp. 105–123). Lexington, MA: Lexington Books.
- Polly, S. (2014, July 2). Workplace well-being is not an oxymoron . Positive Psychology News Daily .
- Quick, C. J. , Quick, J. D. , Nelson, D. L. , & Hurrell, J. J. (2003). Preventive stress management in organizations . Washington, DC: APA.
- Quillian-Wolever, R. E. , & Wolever, M. E. (2003). Stress management at work . In J. C. Quick & L. E. Tetrick (Eds.), Handbook of occupational health psychology (pp. 355–375). Washington, DC: American Psychological Association.
- Ragsdale, J. M. , Beehr, T. A. , Grebner, S. , & Han, K. (2011). An integrated model of weekday stress and weekend recovery of students. International Journal of Stress Management, 18 , 153–180.
- Richardson, K. M. , & Rothstein, H. R. (2008). Effects of occupational stress management intervention programs: A meta-analysis. Journal of Occupational Health Psychology, 13 , 69–93.
- Rodríguez, I. , Kozusznik, M. W. , & Peiró, J. M. (2013). Development and validation of the Valencia Eustress-Distress Appraisal Scale. International Journal of Stress Management, 20 , 279–308.
- Rothbaum, F. , Weisz, J. R. , & Snyder, S. S. (1982). Changing the world and changing the self: A two-process model of perceived control. Journal of Personality and Social Psychology, 42 , 5–37.
- Rotter, J. B. (1966). Generalized expectancies for internal versus external control of reinforcement. Psychological Monographs, 80 , 609.
- Rousseau, V. , & Aubé, C. (2010). Social support at work and affective commitment to the organization: The moderating effect of job resource adequacy and ambient conditions. Journal of Social Psychology, 150 , 321–340.
- Saunders, T. , Driskell, J. E. , Johnston, J. , & Salas, E. (1996). The effect of stress inoculation training on anxiety and performance. Journal of Occupational Health Psychology, 1 , 170–186.
- Schaubroeck, J. , Ganster, D. C. , Sime, W. E. , & Ditman, D. (1993). A field experiment testing supervisory role clarification. Personnel Psychology, 46 , 1–25.
- Selmer, J. (2002). Coping strategies applied by Western vs overseas Chinese business expatriates in China. International Journal of Human Resource Management, 13 , 19–34.
- Siegrist, J. (1996). Adverse health effects of high effort/low reward conditions. Journal of Occupational Health Psychology, 1 , 27–41.
- Siegrist, J. (2010). Effort-reward imbalance at work and cardiovascular diseases. International Journal of Occupational Medicine and Environmental Health, 23 , 279–285.
- Siegrist, J. , Dragano, N. , Nyberg, S. T. , Lunau, T. , Alfredsson, L. , Erbel, R. , et al. (2014). Validating abbreviated measures of effort-reward imbalance at work in European cohort studies: The IPD-Work consortium. International Archives of Occupational and Environmental Health, 87 , 249–256.
- Sinha, B. K. , Willson, L. R. , & Watson, D. C. (2000). Stress and coping among students in India and Canada . Canadian Journal of Behavioural Science/Revue Canadienne Des Sciences Du Comportement , 32 (4), 218–225.
- Sinnott, J. , & Vatz, R. E. (2015, March 13). Maryland doesn’t trust state employees to manage their health . Baltimore Sun .
- Siu, O. L. , Cooper, C. L. , & Phillips, D. R. (2013, July 1). Intervention studies on enhancing work well-being, reducing burnout, and improving recovery experiences among Hong Kong health care workers and teachers . International Journal of Stress Management, 21 , 69–84.
- Sonnentag, S. (2012). Psychological detachment from work during leisure time: The benefits of mentally disengaging from work. Current Directions in Psychological Science, 21 , 114–118.
- Sørensen, O. H. , & Holman, D. (2014). A participative intervention to improve employee well-being in knowledge work jobs: A mixed-methods evaluation study. Work & Stress, 28 , 67–86.
- Spector, P. E. , Chen, P. Y. , & O’Connell, B. J. (2000). A longitudinal study of relations between job stressors and job strains while controlling for prior negative affectivity and strains. Journal of Applied Psychology, 85 , 211–218.
- Spector, P. E. , & Jex, S. M. (1998). Development of four self-report measures of job stressors and strains: Interpersonal Conflict at Work Scale, Organizational Constraints Scale, Quantitative Workload Inventory, and Physical Symptoms Inventory. Journal of Occupational Health Psychology, 3 , 356–367.
- State of Maryland . (n.d.). Wellness program frequently asked questions . Maryland.gov.
- Strentz, T. , & Auerbach, S. M. (1988). Adjustment to the stress of simulated captivity: Effects of emotion-focused versus problem-focused preparation on hostages differing in locus of control . Journal of Personality and Social Psychology, 55 , 652–660.
- Swanberg, J. E. , McKechnie, S. P. , Ojha, M. U. , & James, J. B. (2011). Schedule control, supervisor support and work engagement: A winning combination for workers in hourly jobs? Journal of Vocational Behavior, 79 , 613–624.
- Tarafdar, M. , Tu, Q. , Ragu-Nathan, B. S. , & Ragu-Nathan, T. , (2007). The impact of technostress on role stress and productivity. Journal of Management Information Systems, 24 , 301–328.
- Tepper, B. J. , & Henle, C. A. (2011). A case for recognizing distinctions among constructs that capture interpersonal mistreatment in work organizations . Journal of Organizational Behavior, 32 , 487–498.
- Tetrick, L. E. , & Winslow, C. J. (2015). Workplace stress management interventions and health promotion. Annual Review of Organizational Psychology and Organizational Behavior, 2 , 583–603.
- Thomas, D. C. , Au, K. , & Ravlin, E. C. (2003). Cultural variation and the psychological contract. Journal of Organizational Behavior, 24 , 451–471.
- Toker, S. , Shirom, A. , Melamed, S. , & Armon, G. (2012). Work characteristics as predictors of diabetes incidence among apparently healthy employees. Journal of Occupational Health Psychology, 17 , 259–267.
- Velez, B. L. , Moradi, B. , & Brewster, M. E. (2013). Testing the tenets of minority stres theory in workplace contexts. Journal of Counseling Psychology, 60 , 532–542.
- Verquer, M. L. , Beehr, T. A. , & Wagner, S. H. (2003). A meta-analysis of relations between person–organization fit and work attitudes. Journal of Vocational Behavior, 63 , 473–489.
- Villani, D. , Grassi, A. , Cognetta, C. , Toniolo, D. , Cipresso, P. , & Riva, G. (2013). Self-help stress management training through mobile phones: An experience with oncology nurses. Psychological Services , 10 , 315–322.
- Vischer, J. C. (2007). The effects of the physical environment on job performance: Towards a theoretical model of workspace stress. Stress & Health, 23 , 175–184.
- Wasti, S. A. , & Cortina, L. M. (2002). Coping in context: Sociocultural determinants of responses to sexual harassment . Journal of Personality and Social Psychology , 83, 394–405.
- Willert, M. V. , Thulstrup, A. M. , Hertz, J. , & Bonde, J. P. (2010). Sleep and cognitive failures improved by a three-month stress management intervention . International Journal of Stress Management, 17 , 193–213.
- Xiao Q. , Ottosson I. , & Carlsson I. (2013). Stressors and coping strategies in Chinese and Swedish students at a Swedish university. Chinese Journal of Clinical Psychology, 21 , 309–312.
Related Articles
- Work and Family
- Organizational Climate and Culture
- Physical Activity and Stress Reactivity
- The Roles of Psychological Stress, Physical Activity, and Dietary Modifications on Cardiovascular Health Implications
- Inflammation as a Biomarker Method in Lifespan Developmental Methodology
- Stress and Coping Theory Across the Adult Lifespan
- Occupational Health Psychology
Printed from Oxford Research Encyclopedias, Psychology. Under the terms of the licence agreement, an individual user may print out a single article for personal use (for details see Privacy Policy and Legal Notice).
date: 03 June 2024
- Cookie Policy
- Privacy Policy
- Legal Notice
- Accessibility
- [66.249.64.20|81.177.182.136]
- 81.177.182.136
Character limit 500 /500
Transforming stress through awareness, education and collaboration.
Join the Movement and Give Back!
Generosity gives everyone the power to make a positive change in the lives of others and is a fundamental value anyone can act on., america's # 1 health problem, contemporary stress tends to be more pervasive, persistent and insidious because it stems primarily from psychological rather than from physical threats. it is associated with ingrained and immediate reactions over which we have no control that were originally designed to be beneficial., effects of stress, stress differs for each of us. we respond to stress differently. there are numerous physical as well as emotional responses as illustrated by our list of 50 common signs and symptoms of stress., what is stress, there is no definition of stress that everyone agrees on, what is stressful for one person may be pleasurable or have little effect on others. we all react to stress differently., workplace stress, numerous surveys and studies confirm that occupational pressures and fears are far and away the leading source of stress for american adults and that these have steadily increased over the past few decades., stress management, there is no stress reduction strategy that is a panacea. the key to reducing stress is to prevent it. getting enough sleep, a proper diet, avoiding excess caffeine and other stimulants and taking time out to relax may be helpful in this regard..

People have very different ideas with respect to their definition of stress. Probably the most common is, “physical, mental, or emotional strain or tension.” Another popular definition of stress is, “a condition or feeling experienced when a person perceives that demands exceed the personal and social resources the individual is able to mobilize.”
If life were easy they would have asked for volunteers, stress is ubiquitous and follows us everywhere. since there is simply no avoiding it, the task for mental health professionals has become to further our understanding to improve management in our patients and in ourselves..

To Begin to Manage Your Stress You Must First Identify Your Personal Triggers
Start by taking one of our scientifically validated, AIS certified stress assessments for just $19.95. These tests have been developed to identify your personal stressors and what you can do to control your stress so it doesn’t control you.
In just 15 minutes, this will show you how well you are coping in ten key areas of your life.
The evaluation will also provide a customized report with advice on how you could best manage your stress, plus two valuable free gifts: The Stress Management Journal and the audio A Day Away from Stress.
To master stress, you first need to understand how stress is affecting you physically, emotionally, and socially. The internationally recognized and AIS Certified Stress Mastery Questionnaire (SMQ) provides the information you need to become more aware of not only what to change but how.
Take the SMQ today. It’s easy and quick to take. You will receive your personal 19-page Stress Mastery Report and a complimentary copy of the Stress Mastery Guide and Workbook that will help you understand each of the 11 stress scales and discover new ways to not just reduce but master stress.
Introduction to Stress for Teens
Our newest ais certified free course.
Introduction to Stress for Teens is a free course by The American Institute of Stress to help teenagers understand the stress response and manage every day stressors. In this short course you will learn to understand stress, keep your cool and cope with pressure.
Join the American Institute of Stress

Obtaining credentials from The American Institute of Stress is a designation that sets members apart as stress experts and reflects their commitment to the advancement of innovative and scientifically based stress management protocols.
The AIS Seal and credentials inform the public that the certificate holder commands advanced knowledge of the latest stress research and stress management techniques.
For physicians and other healthcare practitioners it designates your practice as an advanced treatment center for stress related illnesses.
Mismatched: Your Brain Under Stress
This 6-part documentary features some of the world’s leading experts on stress. their collective experience stretches from the first experiments done on the mind/body connection to the latest research into unravelling the unconscious mind. as entertaining as it is informative, mismatched teaches us about what stress is and how to master it., combat stress magazine.

Free magazine for Service Members, Veterans, and First Responders. Edited by Col (Ret) Kathy T. Platoni, PsyD.
Contentment Magazine

Free magazine for everyone. Subscribe to learn about stress and how to manage it in this evidence-based publication.
Finding Contentment Podcast
Finding Contentment focuses on stress and stress-related issues and features interviews with healthcare practitioners and stress experts.
Subscribe today on your favorite podcast app, so you don’t miss any episodes.
Latest Blog Posts
Stress-related news curated by AIS and published weekly
New Research Sheds Light on the Impact of Stress on Tobacco Users

Four Tools For A Healthier Relationship With Stress
A famous quote from Mark Twain says, "Find a job you enjoy doing, and you will never have to work a day [...]

Relax and Unwind: Ways to deal with stress and methods to fall asleep
MARYLAND – Wake up. Work. Repeat. It’s hard to fall asleep when you’re stressed, and day-to-day life can seem mundane, [...]
Shop Our Gift Packs
We assembled a collection of 10 packs of qualified and fun stress management gear. In addition to your own use, these make the perfect gifts for your stressed family members and friends. They will definitely thank you for these useful and thoughtful gift packs.
My Stress Tools The Gift of Relaxation Relaxation-In-A-Box Overcoming Stress Pack Sleep-In-A-Box The Anxiety & Emotional Eating Attack Pack Stress Attack Pack The Chillax Pack Mindfulness Meditation Pack The Unhealthy Behaviors Attack Pack
Online Courses
Learn How to Control Your Reactivity to Stress with our Favorite Certified Online Courses.

Learn to channel your stress for energy
Decrease stress and boost your happiness in just 15 – 20 minutes a day for 21 days with a comprehensive 21 module, online educational program in the practice of stress management.

Free Evidence-based Workforce Resilience and Well-being Course
In this course you will learn behaviors, coping strategies and mindsets that lead to increased resilience against the negative impacts of stress.

Spend the time to learn the best stress reliever there is to enjoy a lifetime of benefits.
An american institute of stress certified course in meditation and more.
Thinking is the cause. Behavior is only the symptom. Behavior change, therefore, is only symptom treatment. We teach you how to think differently to master your stress. This course consists of 13 Modules and 50+ Lessons including PDF lesson downloads and twice monthly group coaching calls. This curriculum includes practical applications in neuroplasticity, evidence-based meditation (that anyone can do!), nutrition, and more.

How can you help?
Giving is not just about donating, it’s about making a difference. Won’t you please donate and help us continue to make a difference?
Our mission is to improve the mental and physical health of the community and the world by setting the standard of excellence of stress management in education, research, clinical care, home life and the workplace. We educate and credential health care professionals and offer products and educational tools for everyone.
Please help us carry out our mission with a donation. The Institute is a 501(c)3 nonprofit corporation. Your tax deductible gift allows us to continue helping you along with Service Members, first responders and civilians navigate stressful situations to have a happier, more rewarding life. Every dollar is greatly appreciated.
The Brain Electric
Watch this free documentary as it takes us across america and the united kingdom to investigate how alpha-stim brain tech is being used by leading researchers and clinicians to normalize your brain without drugs., the body electric, should we accept that medical treatments have side effects evidence suggests that the appropriate use of electroceuticals allows the body to simply take what it needs – creating a powerful effect without side effects. watch this free documentary now., want to receive the latest news and info from ais sign up to receive our enewsletter..
What about our FREE magazines?
Contentment Magazine Combat Stress Magazine
Here’s how you know
- U.S. Department of Health and Human Services
- National Institutes of Health
Let’s Talk About Stress—and Stress Research
Director’s Page Helene M. Langevin, M.D.
July 26, 2022
Stress is a part of life for everyone, and when it comes in short bursts, it’s not necessarily bad. Our natural “fight-or-flight” response can help us mobilize our resources to meet a challenge. But when stress persists (chronic stress), it can lead to both mental and physical health problems. In fact, longstanding evidence from multiple areas of research demonstrates that chronic stress acts like a toxin, permeating our organs and cells and triggering a negative cascade on our hormones, sleep, muscles, metabolism, immune system, and inflammatory responses. And chronic systemic inflammation is emerging as a key factor underlying more than half of all deaths from chronic diseases including heart disease, cancer, diabetes, chronic kidney disease, and nonalcoholic fatty liver.
Chronic stress has long been a focus of research for the National Center for Complementary and Integrative Health (NCCIH), and the interplay between the effects of stress on the mind and the body is an important area of research to me, personally. Many NCCIH-supported studies have looked at the potential role of complementary and integrative health interventions in helping people manage stress and its consequences. Let’s look at one area of research—the effect of stress management interventions on people who are living with a long-term health condition.
With funding from NCCIH, a group of researchers led by Dr. Lori Scott-Sheldon, who was then with The Miriam Hospital in Rhode Island, performed a series of systematic reviews on stress management for people with chronic diseases. The reviews looked at both psychological and physical effects of the interventions, and their findings were promising. For example:
- A review of studies on stress management interventions for adults with heart failure showed improvements in anxiety, depressive symptoms, quality of life, and exercise capacity in people who participated in these interventions.
- A review on mindfulness-based interventions for people living with HIV/AIDS showed reductions in anxiety and depressive symptoms and improved quality of life.
- A review of the evidence on yoga interventions for people living with HIV/AIDS showed improvements in perceived stress, positive affect, and anxiety.
- A review on mindfulness-based interventions for adults with cardiovascular disease showed improvements in anxiety, depression, distress, perceived stress, and systolic blood pressure.
- And finally, a review of yoga interventions for people with type 2 diabetes linked participation in yoga with improvements in measures of glycemic control (HbA1c and both fasting and postprandial [after eating] blood glucose) and in several risk factors for complications of diabetes—blood lipids, body mass index, waist/hip ratio, and cortisol levels (a measure related to the body’s stress response).
The diabetes review doesn’t give us final answers about the value of yoga, though, for two reasons: First, all the studies included in the review were performed outside the United States (most of them in India), and findings obtained in one population may not apply to another. Second, the studies were of only low-to-medium methodological quality, and some important study details were not available.
Fortunately, new research supported by NCCIH may help fill these gaps. Dr. Beth Bock, also of The Miriam Hospital, is leading rigorous NCCIH-funded studies to evaluate the potential role of yoga in diabetes management in U.S. populations, and this work is in progress now. An earlier pilot study showed that the yoga intervention Dr. Bock is testing was highly feasible and acceptable among people with diabetes and that it produced improvements in both psychosocial measures and blood glucose.
Whether or not we face major challenges in our lives the way that people with diabetes or other chronic health problems do, we all need to recognize the importance of stress and the impact it may be having on us. Addressing stress isn’t just about feeling better; it’s about fundamentally promoting health. We can all benefit from learning methods to mitigate stress that have well-documented beneficial effects on inflammation, oxidative stress, stress hormones, blood pressure, blood sugar, and sleep. Some of these tools, such as deep breathing and relaxation techniques, are easy to learn and can help “press reset on stress” —anytime, anywhere. And doing so can have cumulative effects—preventing stress from building up during the day, and giving us a better night’s sleep and more energy the next day for physical activity, which itself helps relieve stress. Think of it as a “positive snowball” effect, leading to better health.
And for those of us in the research community, including NCCIH, learning more about how to prevent, manage, and mitigate stress—and how best to put existing stress management tools into practice—will continue to be a high priority.

Additional Resources
- Stress Fact Sheet
- Feeling Stressed? Ways To Improve Your Well-Being (NIH News in Health)
- Stress Reset Video (The Healthy US Collaborative)

Past Messages From the Director
Disrupting the Disruptors: Convening a Research Discussion To Mitigate Endocrine-Disrupting Chemicals May 13, 2024
NCCIH Heads to Cleveland for ICIMH April 5, 2024
Addressing Pain Requires Understanding How We Heal March 27, 2024
More messages
Watch CBS News
Why "stress bragging" can annoy your co-workers and hurt your career
By Megan Cerullo
Edited By Alain Sherter
May 28, 2024 / 3:13 PM EDT / CBS News
Constantly announcing to co-workers how swamped you are at the office isn't likely to impress your colleagues — in fact, it may have the opposite effect, new research shows.
"Stress bragging" or "busy bragging" about your overflowing plate often leads to resentment from peers, a study from the University of Georgia (UGA) shows. It also tends to make boasters appear less competent at their job, the researchers found.
"This is a behavior we've all seen, and we all might be guilty of at some point," Jessica Rodell, a professor of management at UGA's Terry College of Business and the study's lead author, told UGA Today , an online newspaper geared to the university. "When I was wondering about why people do this, I thought maybe we are talking about our stress because we want to prove we're good enough. We found out that often backfires."
Despite the adage, "If you want something done, ask a busy person" (variously attributed to Benjamin Franklin or writer Elbert Hubbard), the findings suggest that harping on about your busy schedule is unlikely to engender much in the way of good will at work.
To assess such attitudes, the UGA researchers asked hundreds of subjects to rate a fictitious co-worker who made statements about an imaginary conference, including that it was "just one more thing on my full plate. And I was already stressed to the max…you have no idea the stress that I am under."
Study respondents said they found that type of individual to be less likable and less competent than a colleague who simply said work had been stressful or, alternatively, who had positive things to say about the conference. Participants also said they wouldn't be inclined to lend a complaining coworker a helping hand. Researchers also studied real-life workplace braggarts and found their colleagues often perceived them negatively.
"People are harming themselves by doing this thing they think is going to make them look better to their colleagues," Rodell said.
No badge of honor
Meanwhile, "stress bragging" perpetuates an unhealthy notion, according to the study's authors — that work should be stressful, and by extension anyone who isn't sweating their job isn't cutting the mustard.
"When somebody is constantly talking about and bragging about their stress, it makes it seem like it is a good thing to be stressed," Rodell told UGA Today . "It just spills over onto the co-worker next to them. They wind up feeling more stressed, which leads to higher burnout or withdrawal from their work. Think of it as this spiraling contagious effect from one person to the next."
Of course, all workers may experience stress of one kind or another, and the study doesn't suggest that people should mask their emotions. And co-workers who were perceived as being genuinely busy didn't invite ill will, the research found.
"If you genuinely feel stressed, it's OK to find the right confidant to share with and talk about it," she said. "But be mindful that it is not a badge of honor to be bragged about — that will backfire," Rodell said.
Megan Cerullo is a New York-based reporter for CBS MoneyWatch covering small business, workplace, health care, consumer spending and personal finance topics. She regularly appears on CBS News 24/7 to discuss her reporting.
More from CBS News

Mediterranean diet associated with 23% reduction in mortality, study finds

Man accused of killing Georgia nursing student pleads not guilty

Trump lawyer eager to move forward with appeal of New York conviction

Trump may face travel restrictions in some countries after conviction
- Alzheimer's disease & dementia
- Arthritis & Rheumatism
- Attention deficit disorders
- Autism spectrum disorders
- Biomedical technology
- Diseases, Conditions, Syndromes
- Endocrinology & Metabolism
- Gastroenterology
- Gerontology & Geriatrics
- Health informatics
- Inflammatory disorders
- Medical economics
- Medical research
- Medications
- Neuroscience
- Obstetrics & gynaecology
- Oncology & Cancer
- Ophthalmology
- Overweight & Obesity
- Parkinson's & Movement disorders
- Psychology & Psychiatry
- Radiology & Imaging
- Sleep disorders
- Sports medicine & Kinesiology
- Vaccination
- Breast cancer
- Cardiovascular disease
- Chronic obstructive pulmonary disease
- Colon cancer
- Coronary artery disease
- Heart attack
- Heart disease
- High blood pressure
- Kidney disease
- Lung cancer
- Multiple sclerosis
- Myocardial infarction
- Ovarian cancer
- Post traumatic stress disorder
- Rheumatoid arthritis
- Schizophrenia
- Skin cancer
- Type 2 diabetes
- Full List »
share this!
May 28, 2024
This article has been reviewed according to Science X's editorial process and policies . Editors have highlighted the following attributes while ensuring the content's credibility:
fact-checked
peer-reviewed publication
trusted source
Sleep study finds post-karaoke stress is strengthened by REM sleep
by Society for Neuroscience

We're all familiar with the fact that the quality of our sleep has a strong impact on our waking lives. But what you may not know is that sleep stages contribute to how we process life experiences and memories in different ways. Deep sleep, or slow wave sleep (SWS), is associated with memory consolidation, which makes memories longer lasting.
Our understanding of rapid eye movement (REM) sleep is less clear: although it is associated with the way we process emotions and how well we remember more emotionally charged experiences, there is conflicting evidence for the exact role it plays.
In a study led by Anu-Katriina Pesonen from the SleepWell Research Program Unit at the University of Helsinki in Finland, the scientists sought to shed more light on the role of sleep stages in emotional processing by investigating how SWS and REM sleep affect stress-related responses. The paper is published in the journal eNeuro .
They divided 29 participants into two groups: one in which SWS was selectively disrupted, and one in which REM was disrupted. To trigger stress responses in participants, experimenters applied a karaoke paradigm in which participants performed karaoke without being able to listen to their singing and then later had to listen to clips of their recordings with no background music.
Physiological stress responses induced by the clips were measured before the night of sleep in which disruption occurred and in two separate mornings after the night of sleep disruption.
The scientists found that suppressing SWS while keeping REM sleep resulted in stronger physiological stress responses the next morning than those following suppression of REM sleep. REM sleep's preservation of the stress response may be due to its strengthening of how emotionally charged a stressful memory is.
This study advances our understanding of how sleep stages differentially impact emotional responses and may inform treatment options for anxiety disorders.
Explore further
Feedback to editors

Cancer patients often do better with less intensive treatment, new research finds
7 hours ago

Study sheds new light on the contribution of dopamine to reinforcement learning
13 hours ago

Almost 1 in 3 Americans know someone who's died from a drug overdose
Jun 1, 2024

Antibody discovery promises new hope in influenza B battle, paves way for universal vaccine

Eye-tracking techniques could help primary care providers diagnose autism sooner, more accurately
May 31, 2024

This self-powered sensor could make MRIs more efficient

Not eating can hinder weight loss, study in fruit flies suggests

Neuroscience research suggests ketones can enhance cognitive function and protect brain networks

New research finds antidepressants may help deliver other drugs into the brain

Mediterranean diet tied to one-fifth lower risk of death in women
Related stories.

Dreaming is linked to improved memory consolidation and emotion regulation
May 14, 2024

Research shows how stress activates neurons that disrupt sleep
Dec 13, 2023

Why do embarrassing memories seem to surface at night?
Jun 27, 2022

Brain wave activity associated with circadian preferences
Nov 3, 2017

Sleep disorders in parents and children associated with more parental stress, study finds
Jan 25, 2023

Sleep deprivation makes us less happy and more anxious, says review of more than 50 years of research
Dec 21, 2023
Recommended for you

Researchers identify a genetic cause of intellectual disability affecting tens of thousands

Mobile app effective in smoking cessation, triples chance of success

Infants hear significantly more speech than music at home, study finds
Let us know if there is a problem with our content.
Use this form if you have come across a typo, inaccuracy or would like to send an edit request for the content on this page. For general inquiries, please use our contact form . For general feedback, use the public comments section below (please adhere to guidelines ).
Please select the most appropriate category to facilitate processing of your request
Thank you for taking time to provide your feedback to the editors.
Your feedback is important to us. However, we do not guarantee individual replies due to the high volume of messages.
E-mail the story
Your email address is used only to let the recipient know who sent the email. Neither your address nor the recipient's address will be used for any other purpose. The information you enter will appear in your e-mail message and is not retained by Medical Xpress in any form.
Newsletter sign up
Get weekly and/or daily updates delivered to your inbox. You can unsubscribe at any time and we'll never share your details to third parties.
More information Privacy policy
Donate and enjoy an ad-free experience
We keep our content available to everyone. Consider supporting Science X's mission by getting a premium account.
E-mail newsletter
Suggestions or feedback?
MIT News | Massachusetts Institute of Technology
- Machine learning
- Social justice
- Black holes
- Classes and programs
Departments
- Aeronautics and Astronautics
- Brain and Cognitive Sciences
- Architecture
- Political Science
- Mechanical Engineering
Centers, Labs, & Programs
- Abdul Latif Jameel Poverty Action Lab (J-PAL)
- Picower Institute for Learning and Memory
- Lincoln Laboratory
- School of Architecture + Planning
- School of Engineering
- School of Humanities, Arts, and Social Sciences
- Sloan School of Management
- School of Science
- MIT Schwarzman College of Computing
Scientists identify mechanism behind drug resistance in malaria parasite
Press contact :.

Previous image Next image
Share this news article on:
Related links.
- Peter Dedon
- Antimicrobial Resistance Interdisciplinary Research Group
- Singapore-MIT Alliance for Research and Technology (SMART)
- Department of Biological Engineering
Related Topics
- Drug resistance
- Antibiotics
- Biological engineering
- Drug development
- International initiatives
- Collaboration
Related Articles

Satellite-based method measures carbon in peat bogs

Newly discovered bacterial communication system aids antimicrobial resistance

SMART launches research group to advance AI, automation, and the future of work

A novel combination therapy counters antibiotic-resistant Mycobacterium abscessus infections
Previous item Next item
More MIT News

MIT Corporation elects 10 term members, two life members
Read full story →

Diane Hoskins ’79: How going off-track can lead new SA+P graduates to become integrators of ideas

Chancellor Melissa Nobles’ address to MIT’s undergraduate Class of 2024

Noubar Afeyan PhD ’87 gives new MIT graduates a special assignment

President Sally Kornbluth’s charge to the Class of 2024

Commencement address by Noubar Afeyan PhD ’87
- More news on MIT News homepage →
Massachusetts Institute of Technology 77 Massachusetts Avenue, Cambridge, MA, USA
- Map (opens in new window)
- Events (opens in new window)
- People (opens in new window)
- Careers (opens in new window)
- Accessibility
- Social Media Hub
- MIT on Facebook
- MIT on YouTube
- MIT on Instagram
- Open access
- Published: 31 May 2024
Genome-wide identification and expression analysis of the SPL transcription factor family and its response to abiotic stress in Pisum sativum L
- Long Li 1 , 3 ,
- Jian bo Xu 2 ,
- Zhi wen Zhu 2 ,
- Xiao zong Wu 2 , 5 &
- Yu ke Geng 1
BMC Genomics volume 25 , Article number: 539 ( 2024 ) Cite this article
21 Accesses
Metrics details
Squamous promoter binding protein-like ( SPL ) genes encode plant-specific transcription factors (TFs) that play essential roles in modulating plant growth, development, and stress response. Pea ( Pisum sativum L. ) is a coarse grain crop of great importance in food production, biodiversity conservation and molecular genetic research, providing genetic information and nutritional resources for improving agricultural production and promoting human health. However, only limited researches on the structure and functions of SPL genes exist in pea ( PsSPLs ). In this study, we identified 22 PsSPLs and conducted a genome-wide analysis of their physical characteristics, chromosome distribution, gene structure, phylogenetic evolution and gene expression patterns. As a result, the PsSPLs were unevenly distributed on the seven chromosomes of pea and harbored the SBP domain, which is composed of approximately 76 amino acid residues. The phylogenetic analysis revealed that the PsSPLs clustered into eight subfamilies and showed high homology with SPL genes in soybean. Further analysis showed the presence of segmental duplications in the PsSPLs . The expression patterns of 22 PsSPLs at different tissues, developmental stages and under various stimulus conditions were evaluated by qRT-PCR method. It was found that the expression patterns of PsSPLs from the same subfamily were similar in different tissues, the transcripts of most PsSPLs reached the maximum peak value at 14 days after anthesis in the pod. Abiotic stresses can cause significantly up-regulated PsSPL19 expression with spatiotemporal specificity, in addition, four plant hormones can cause the up-regulated expression of most PsSPLs including PsSPL19 in a time-dependent manner. Therefore, PsSPL19 could be a key candidate gene for signal transduction during pea growth and development, pod formation, abiotic stress and plant hormone response. Our findings should provide insights for the elucidating of development regulation mechanism and breeding for resistance to abiotic stress pea.
Peer Review reports
Pea ( Pisum sativum L. ) is a legume plant and a member of the genus Pisum . It is an annual climbing plant and an important source of food [ 1 , 2 ]. Pisum sativum is a source of high-quality proteins and other nutrients such as dietary fiber, vitamin C, vitamin K, and minerals [ 3 , 4 ]. Additionally, pea is used to produce green manure, which improves soil quality by fixing nitrogen and providing nutrients to crops [ 5 , 6 ]. Meanwhile, the strong drought - and cold - resistant properties made pea adaptable to a wide range of climates [ 7 ].
Transcription factors (TFs) are a class of proteins that regulate gene transcription. They regulate gene expression by binding to specific regions in gene promoters, activating or inhibiting gene transcription, and thus regulate a variety of biological processes, such as plant growth, development, differentiation and response to environmental stimuli [ 8 , 9 ]. Squamosa promoter binding protein-like (SPL) TFs are a ubiquitous family of TFs in various plants and contain a conserved SBP domain, two Cys2/His2 type zinc fingers, and two types of zinc fingers which is a structurally stable zinc finger motif formed by two cysteines (Cys) and two histidines (His) via ring and helical connections, and this domain binds to specific regions of DNA [ 10 , 11 , 12 ]. SPL is usually involved in plant growth, flower organ development and fruit development [ 10 , 13 ]. In order to explore the biological functions and regulatory mechanisms of SPLs in plant growth, development and stress response, various plant species have been studied and elaborated by scholars, such as Arabidopsis thaliana [ 14 , 15 ], Morus alba [ 16 ], Nicotiana tabacum [ 17 ], Zea mays [ 18 ], Setaria italica [ 19 ], Vaccinium corymbosum [ 20 ], Vitis vinifera [ 21 ], Fagopyrum tataricum [ 22 ], Solanum lycopersicum [ 23 ], Codonopsis pilosula [ 23 ], Triticum aestivum [ 24 ], and Jatropha curcas [ 25 ].
SPL TFs were first identified in Snapdragon ( Antirrhinum majus L .). AmSPL regulates the formation and development of flower organs via binding to promoters of genes related to flower development [ 26 ]. AtSPL9 and AtSPL15 in Arabidopsis thaliana are highly expressed in stem apex meristem and participate in leaf differentiation to regulate plant leaf morphology and structure [ 27 , 28 ]. Cui et al. confirmed that AtSPL9 is negatively regulated by miR156, which promotes early flowering of Arabidopsis thaliana under salt and drought stress. In addition, AtSPL9 can compete with TT8 (Transparent testa8) combined with PRODUCTION OFANTHOCYANIN PIGMENT 1 (PAP1) [ 29 ], such competition interferes with the stability of MYB-bHLH-WD40 (MBW) transcription-activating complex, directly prevents the expression of flavonoid synthesis gene dihydroflavonol 4-reductase (DFR), and thus negatively regulates anthocyanin accumulation [ 15 ]. AtSPL3 and AtSPL4 are significantly differentially expressed in the apical meristem of Arabidopsis thaliana , but both are involved in the regulation of flowering [ 15 , 30 ]. Loss-of-function mutants in OsSPL14 , OsSPL16 and OsSPL18 lead to flower dysplasia, including increased inflorescence branching, atypical flower morphology and anther development defects [ 31 ]. This implies that these SPL TFs play essential regulatory roles in the formation and development of rice flower organs.
In this study, we identified PsSPLs from pea genome sequence obtained from publicly available databases using bioinformatics analysis. A genome-level comprehensive analysis was performed to explore their structural characteristics, motif composition, chromosome localization and evolutionary relationships. Then, we used real-time quantitative fluorescent PCR (qRT-PCR) to analyze the expression patterns of PsSPLs of different tissues, expression changes during pod formation stage, induced expression level under abiotic stress and hormone response, separately. The results provide an experimental basis for further revealing the mechanism of pea SPL gene family in regulating the growth and development, yield and environmental response, which possesses important application value for molecular breeding of pea with high yield and quality.
Gene identification
The whole-genome sequence of Pisum sativum was retrieved from the Ensembl Genomes website ( http://ensemblgenomes.org/ ), whereas the whole-genome sequence of Arabidopsis thaliana was downloaded from The Arabidopsis Information Resource (TAIR) website ( https://www.arabidopsis.org/ ). BLASTp [ 32 ](with a score cutoff of ≥ 100 and an e-value cutoff of ≤ 1e-10) was used to align AtSPL with the complete genome sequence of closely related species rye to identify candidate SPL genes. Subsequently, the hidden Markov model (HMM) of the SBP conserved domain was retrieved from the Pfam database ( http://pfam.sanger.ac.uk/ ), and HMMER 3 ( http://plants.ensembl ) was used to identify non-redundant SPL genes [ 33 ]. The selected SPL genes were confirmed using the SMART tool ( http://smart.embl-heidelberg.de/ ) [ 34 , 35 ]. A total of 22 SPL genes were obtained for the subsequent analyses. ExPasy tool was used to determine the physical characteristics of the genes, including sequence length, isoelectric point (pI), and molecular weight (MW). The WoLFPSORT tool ( https://wolfpsort.hgc.jp/ ) was used to predict the subcellular localization of the selected SPL genes.
Gene structure, chromosomal distribution and gene duplication analysis and classification of the PsSPLs
The MEME website ( http:/meme.nbcr.net/meme/intro.html ) was used to identify the motifs in PsSPLs , with a maximum of 10 motifs set for the search. The GSDS ( http://gsds.cbi.pku.edu.cn ) tool was used to analyze the distribution of introns and exons in the sequences of PsSPLs . The Circos website was used to identify the location of PsSPLs in chromosomes. MCScanX toolkit was used to evaluate the collinear duplication events on the chromosomes [ 36 ]. Mega7.0 software was used to perform multiple sequence comparison of SPL genes from pea and Arabidopsis thaliana , and the phylogenetic analysis of different SPL genes was performed by neighborhood linkage (NJ) method and phylogenetic tree construction [ 37 ]. At the same time, the NJ method was also used for phylogenetic analysis of SPL genes from Pea, Oryza sativa , Zea mays , Hordeum vulgare , Arabidopsis thaliana , Triticum aestivum (hexaploid) and Aegilops tauschii Coss , Moreover, the evolutionary relationships among different species and subfamilies are further discussed.
Evaluation of varying growth conditions and the effect of different hormone treatments on Pisum sativum
In this study, pea No. 6 cultivar provided by Zhengzhou University of Light Industry was taken as the research material. Fully mature pea seeds were selected and evenly sown in a greenhouse under the following growth conditions: 16 h light at 28 °C / 8 h darkness at 20 °C, and 75% constant relative humidity. The Tissues (roots, stems, leaves, anthers and styles, seeds) of pea plants with the same growth state at 21-day growth are collected and stored in an ultra-low temperature refrigerator of -80 °C. Parallelly, Pea plants were treated with four abiotic stresses, including salt stress (5% sodium chloride), low temperature stress (4 °C), drought stress (30% PEG 6000) and high temperature stress (40 °C) for 0, 1, 4 and 12 h, respectively, and the untreated plants obtained at the same time point were used as controls, the whole plants were collected and stored in an ultra-low temperature. After that, the pea plants at the filling stage were treated with ABA, IAA, JA, SA and GA3 at concentrations of 100µM for 0, 1, 4 and 12 h, respectively and the whole plants were sampled and preserved at ultra-low temperature. In the above experiments, three plants were selected for one type of treatment, and three independent biological replicates were set up for each.
Total RNA extraction, cDNA synthesis, and qRT-PCR analysis
RNA extraction kit (Tiangen Biotech, Beijing, China) was used to extract RNA from different tissue samples collected. HiScript Q RT Super Mix for qPCR (Vazyme #R122) was used for reverse transcription to obtain cDNA, and AceQ qPCR SYBR Green Master Mix was used for qRT-PCR analysis of SPL gene. Primers for SPL gene qRT-PCR experiments were designed using Primer5.0 software (Additional file 5: Table S5 ). In this study, the glyceraldehyde-3-phosphate dehydrogenase (GADPH) gene was used as an internal reference for gene expression quantification. The experiment was repeated three times and the relative gene expression was calculated by 2^-(ΔΔCt) method. Compared with the control group, we defined a significant difference in expression levels as ≥ 2-fold or ≤ 0.5-fold [ 38 ].
Statistical analysis
In this study, all data were subjected to analysis of variance (ANOVA) using the JMP 6.0 software (SAS Institute). The least significant difference (LSD) test was performed to determine the significance of differences between different treatment groups [ 38 , 39 ]. A significance level (e.g., 0.05 or 0.01) is typically set in the LSD test to determine if the differences are statistically significant. Origin version 8.0 (OriginLab, Northampton, MA, USA) was used to generate the histograms for gene expression levels.
Identification of SPL genes in Pea
A comprehensive and systematic analysis of the PsSPLs was conducted, leading to identification and characterization of 22 PsSPLs . The length of proteins encoded by these genes ranged from 139 (PsSPL3) to 1025 (PsSPL9) amino acids. The maximum molecular weight of the proteins encoded by PsSPL was 113.918 kDa ( PsSPL18 ), whereas the minimum molecular weight was15.9 kDa ( PsSPL4 ). The pI values ranged from 6 to 9.28 ( PsSPL7 and PsSPL12 ), with an average value of 7.622. All the PsSPLs harbored the conserved SBP domain, and four of them contain ANK (ankyrin) domain (Additional file 1: Table S1 ). The ANK domain is a conserved domain involved in protein-protein interaction and it plays an important role in various biological processes, including cell signaling, protein interactions and assembly of protein complexes. Subcellular localization prediction revealed that all the PsSPLs were localized in the nucleus, however. some were observed in different cellular tissues with 9 SPLs localized in the endoplasmic reticulum, 6 SPLs in the chloroplasts and seven in the cytoplasm (Additional file 1: Table S1 ).
PsSPLs exhibit high evolutionary conservation
Phylogenetic analysis of 22 PsSPLs from pea and 16 AtSPLs from Arabidopsis thaliana was conducted in this study. The PsSPLs formed three branches (Group 1–8) based on the classification method proposed by Cenci and Rouard consistent with the classification of SPL proteins in Arabidopsis thaliana [ 40 ]. This finding indicated conservation of SPLs across the evolutionary process without any loss events. In addition, we observed an uneven distribution of SPLs exist among the eight subfamilies. Subfamilies II and VI contained the largest number of members (4 PsSPLs ), whereas subfamily III possessed the lowest number of genes (only 1 PsSPL ) (Fig. 1 ). Subfamilies VII, VIII, and IV owned the same number of SPL s (3 PsSPLs ). Subfamilies I and V each contain 2 PsSPLs (Fig. 1 ; Additional file 1: Table S1 ). Comparative analysis of the phylogenetic tree revealed that PsSPL clustered with AtSPL (bootstrap support ≥ 70), implying that these proteins may exert similar biological functions. These findings would provide a basis for further exploring the evolution and function of the SPL gene family in plants.

Phylogenetic analysis of SPL proteins in pea and Arabidopsis thaliana . 22 PsSPLs were divided into eight clades (I–IX) and indicated with different colors. The red pentacle represents Pea, the black circle represents Arabidopsis thaliana
Conserved motifs and structure analysis of the PsSPLs
Comparing the exons and introns of genes provided a comprehensive understanding of gene structure, function, and regulation in an organism [ 41 , 42 ]. The results showed that different genes exhibited varying numbers of exons and introns, ranging from 0 to 14. PsSPL19 exhibited the highest number of introns (14), whereas PsSPL5 lacked intronic structures (Fig. 2 ). PsSPL1 , PsSPL7 , PsSPL18 , and PsSPL19 , members of subfamily II, exhibited the most complex gene structures, indicating that subfamily II may have different functions. Typically, members of the same gene family exhibit conserved intron and exon structures. This conservation can be attributed to the conserved structure of introns and exons caused by gene replication and recombination during the evolution of gene family members.
We identified 10 conserved motifs conserved across the genomic sequences of different genes using the MEME tool [ 43 , 44 ], indicating the homology and conservation of regions in these genes. We named these motifs as Motif 1–10. The different subfamilies exhibited some differences in motifs, but all genes had Motif 1, indicating that Motif 1 is a conserved domain in SPL genes (Fig. 2 ). Subfamily II exhibited 9 of the 10 motifs identified in this study (lacked Motif 10). All genes exhibited Motif 1, Motif 2, and Motif 3. Subfamilies IV, V, VII, and VIII had Motif 10, which was absent in the other subfamilies. Motif 2 was located at the beginning of all patterns, whereas Motif 1 was positioned between Motif 2 and Motif 3 (Fig. 2 ; Additional file 2: Table S2 ). The conservation of motifs implied that the subfamily potentially emerged through evolutionary processes such as gene duplication and recombination. These findings indicate that the genes may form clusters in the genome with similar structural and functional features.

Phylogenetic relationships, gene structure, and motif distributions of PsSPL genes. Phylogenetic tree was constructed for each node with 1000 replicates using the NJ method ( A ). Exons and introns are indicated by yellow rectangles and grey lines, respectively ( B ). These numbers indicate the annotation file of the different phases of gene CDS, which are defined as “0”, “1”, and “2”. C Amino acid motifs in the SPL proteins (1–10) were represented by colored boxes, the black lines indicate relative protein lengths
Chromosomal distribution and gene duplication of PsSPLs
The SPL genes were mapped to different chromosomes using the latest version of the Pea genome database [ 45 ]. The 22 PsSPLs were unevenly distributed across seven chromosomes (Fig. 3 ). Chr1 and Chr3 harbored the highest number of PsSPL s (4 genes, approximately 18.18%), followed by Chr4, Chr5, Chr6, and Chr7, which each contained three SPLs (approximately 13.63% each). Chr2 harbored the least number of PsSPLs (2 genes, approximately 9.09%). The PsSPLs are randomly distributed across different chromosomes with no significant correlation between the distribution of SPLs on chromosomes to their function and structure.
Tandem duplication events and segmental duplication events are key forms of gene duplication processes, which contribute to genomic diversity and play important roles in shaping the structure and function of the genome [ 46 , 47 ]. In this study, no any tandem duplication event was detected on the chromosomes (Fig. 4 ; Additional file 3: Table S3 ). However, we found eight homologous sequences produced by four pairs of fragments distributed on five chromosomes, this indicated the existence of evolutionary relationships among PsSPLs , Chr5 contains the largest number of PsSPLs ( n = 3), followed by Chr 7 ( n = 2), while Chr 3, Chr 4, and Chr 6 each contains only one PsSPL (Fig. 4 ).

Schematic representation of the chromosomal distribution of PsSPLs . Vertical bars represent the chromosomes of Pea. The chromosome number is indicated to the left of each chromosome. The scale on the left represents chromosome length

Schematic representation of the chromosomal distribution and interchromosomal relationships of PsSPLs . Colored lines denote all synteny blocks in the Pea genome, and the red lines denote duplicated SPL pairs. The chromosome number is denoted at the bottom of each chromosome
PsSPLs are evolutionarily related to SPLs in other species
We compared the PsSPL gene with SPL genes from two dicot plants ( Arabidopsis thaliana and Glycine max ) and four monocot plants ( Setaria italica , Triticum aestivum , Oryza sativa and Chenopodium quinoa ) to evaluate the genomic diversity among different species and explore the common ancestors and evolutionary relationships. We constructed a phylogenetic tree using the NJ method based on 10 conserved motifs identified in the genes, obvious distinctions in motifs among the different species were observed, but the same subfamily exhibited motif conservation (Fig. 5 ). Most genes harbored Motif 1, Motif 2, and Motif 4. Subfamily II exhibited all the 10 motifs, implying that this subfamily may have a more complex and diverse gene regulatory network compared with the other subfamilies. All members of the subfamily except for subfamily VI started with Motif7. Most subfamilies exhibited high homology with the Gm SPL gene family of soybean, indicating that pea and soybean may share a common ancestor or had high degree of structural and sequence conservation during evolution.

Phylogenetic relationships and motif compositions of the PsSPLs with six different plant species ( Arabidopsis thaliana , Solanum lycopersicum , Vitis vinifera , Sorghum bicolor Moench , Oryza sativa and Zea mays ). Outer panel: an unrooted phylogenetic tree constructed using Geneious R11 with the neighbor-joining method. Inner panel: distribution of conserved motifs in SPL proteins. The differently colored boxes represent different motifs and their positions in each SPL protein sequence. The sequence information for each motif is provided in Additional file 2 (Table S2 )
We conducted a collinearity analysis based on the genes of Pisum sativum and six representative species to elucidate the evolutionary relationships and functional conservation across different species. The results revealed collinearity between 22 PsSPL genes and genes from A rabidopsis thaliana (15), Setaria italica (18), Chenopodium quinoa (23), Glycine max (29), Oryza sativa (29), and Triticum aestivum (56). Monocot plants and pea exhibited a closer evolutionary relationship than the relationship between dicot and monocot plants. We identified 52 gene pairs shared between Glycine max and Pisum sativum , 15 gene pairs shared with Arabidopsis thaliana , 7 gene pairs shared with Oryza sativa , 7 gene pairs shared with Setaria italica , 6 gene pairs shared with Triticum aestivum , and 6 gene pairs shared with Chenopodium quinoa .
Comparative analysis of collinearity among the six plants revealed that the number of collinear SPL genes between Pisum sativum and Triticum aestivum and Chenopodium quinoa was the lowest, whereas the number of collinear SPL genes between Pisum sativum and Glycine max was the highest. Interestingly, these species shared some common collinear genes. For example, PsSPL3 exhibited collinearity with AT5G18830.3 / Os05t0408200-01 / TraesCS1A02G255300.1 / KQL15519. This finding indicates that these collinear genes are highly conserved and possibly existed before the divergence of these species (Fig. 6 ; Additional file 4: Table S4 ). Collinear genes are used to explore genomic similarities and differences, evolutionary relationships, and functional conservation among different species. The findings on collinear genes provide information on genetic variations and evolutionary relationships among different species.

Synteny analyses of the SPL genes between Pisum sativum and six representative plant species ( Arabidopsis thaliana , Glycine max , Chenopodium quinoa , Oryza sativa, Triticum aestivum and Setaria italica ). Gray lines on the background indicate the collinear blocks in Pisum sativum and other plant genomes; red lines highlight the syntenic of SPL gene pairs in Pisum sativum
Expression patterns of the PsSPL s in different plant organs
In order to evaluate the potential function of PsSPLs , qRT-PCR was used to analyze the expression of 22 PsSPLs in four organs: root, stem, leaf, flower and fruit. The results show that PsSPLs were expressed differently in these four organs, which reflects the specific biological functions and adaptation strategies of plants at different growth and development stages and under different environmental conditions. Three genes ( PsSPL3 , PsSPL7 , and PsSPL19 ) were most expressed in stems, while eight genes ( PsSPL2 , PsSPL5 , PsSPL6 , PsSPL9 , PsSPL11 , PsSPL14 , PsSPL15 , and PsSPL20 ) were most expressed in leaves. PsSPL1 , PsSPL12 , PsSPL18 and PsSPL20 were highly expressed in flowers (Fig. 7 ). Genes from the same subfamily may have maintained similar expression patterns over the course of evolution, reflecting that they may share a common ancestor and have maintained similar regulatory patterns over the course of evolution. It is obvious that the expression levels of all PsSPLs in roots are lower than those in stems, leaves and flowers, so we speculate that SPLs may be closely related to the development of stems, leaves and flowers of plants.
Previous studies have shown that the expression level and pattern of SPL during fruit development may be related to biological processes such as fruit growth, ripening and storage. Therefore, we selected 15 representative genes for qRT-PCR verification at five post-anthesis stages (7DPA, 14DPA, 21DPA, 28DPA and 35DPA). As shown in Fig. 7 , almost all genes showed differential expression patterns at different time periods. In pea fruits, we found that PsSPL4 had the most significant differential expression at 14DPA stage, while PsSPL3 showed a downward trend with the increase of time. Interestingly, PsSPL9 remained stable over time, suggesting that it may not be associated with fruit development. In addition, we found that most of the significantly differentially expressed genes (DEGs) were concentrated at 14DPA stage, such as PsSPL2 , PsSPL4 , PsSPL6 , PsSPL8 , PsSPL11 , PsSPL12 , PsSPL13 , PsSPL14 , PsSPL15 , PsSPL17 , PsSPL2 , PsSPL4 , PsSPL6 , PsSPL8 . PsSPL19 and PsSPL22 . These results indicated that the expression of most PsSPLs were mainly concentrated in the early stage of pea fruit. The expression level of PsSPL1 reached its maximum value at 35DPA stage, which may be related to the later stage of fruit development. In addition, we found that the expression of PsSPL7 was the highest in the pod, and most of the genes showed significant differential expression. The expression of PsSPL5 and PsSPL16 was not significantly different at different times, indicating that it was not related to pod development of pea. The heat map showed that there was a correlation between different PsSPL s, and most PsSPLs were positively correlated. However, some PsSPLs were negatively correlated, such as PsSPL6 and PsSPL21 / PsSPL1 , and PsSPL1 and PsSPL9 ( P < 0.05).

Tissue-specific gene expression of 22 PsSPLs and gene expression during fruit development. Expression patterns of 22 PsSPLs and in fruit of different stage and flower, leaf, root, stem were detected by qPCR. Error bars are obtained from three measurements. Lowercase letters indicate significant differences between treatments ( α = 0.05, LSD) ( A , C , E ). Coexpression analysis of 22 PsSPLs ( B , D , F )
Expression patterns of PsSPLs under various abiotic stress conditions
Prediction of gene function by exposing plants to various stress treatments can offer valuable insights into elucidating the biological functions of genes under adverse conditions, providing information on plant adaptation mechanisms and regulatory networks in response to stress [ 48 ]. We used four stress treatments to explore the effects of various stress conditions on the expression patterns of PsSPL genes in different tissues. The results showed that various stress conditions induced differential expression of PsSPL genes in different tissues and at different time points. PsSPL genes are implicated in stress signaling pathways, so their expression levels are upregulated with increase in treatment time, indicating their important regulatory role in alleviating stress in plants. The expression of most SPL genes was upregulated in stems under cold stress conditions. PsSPL1 , PsSPL3 , PsSPL7 , PsSPL10 , PsSPL18 , and PsSPL19 genes exhibited significant differential expression in roots, leaves, and stems, with upregulation at the early stages of stress treatment and then downregulation at the late stages. PsSPL19 had the highest expression level under PEG and salt stress condition, and the expression of the other genes was also upregulated under these two stress treatments. Conversely, the expression of PsSPL5 , PsSPL15 , PsSPL16 , PsSPL17 , and PsSPL20 were downregulated under PEG and salt stress conditions. Tissue-specific responses were mainly observed in stems under cold and heat stress treatments. PsSPL4 , PsSPL10 , PsSPL13 , PsSPL14 and PsSPL18 genes displayed similar expression patterns, with increases expression levels in different tissues observed over treatment time. On the contrary, the expression of PsSPL5 , PsSP16 and PsSPL20 was downregulated over time. The genes showed different expression patterns under different treatments, and the significant downregulation observed at the start of stress induction may be attributed to the rapid regulation and adaptation process of genes to the treatments. PsSPL19 expression was upregulated under the four stress treatments in different tissues, indicating its potentially important role in stress responses. We performed correlation analysis and generated a heatmap to show the correlation in expression levels among PsSPL members under stress. The results showed that the expression levels of PsSPL2 , PsSPL3 , PsSPL6 , PsSPL9 , PsSPL11 , PsSPL12 , PsSPL14 PsSPL15 , PsSPL18 , and PsSPL19 , genes were significantly positively correlated, whereas the expression of PsSPL1 was significantly positively correlated with PsSPL5 expression (Fig. 8 ). The expression of some PsSPL genes was significantly negatively correlated with the expression of PsSPL5 and PsSPL20 .

Expression of PsSPLs in plants subjected to abiotic stresses (PEG, NaCl, heat, and cool treatments) at the seedling stage in three organs (root, stem, and leaf). ( A ) Changes in expression of representative genes analyzed by qRT-PCR. Error bars were obtained from three measurements. The lowercase letter above the bar indicates a significant difference (α = 0.05, LSD) among the treatments. ( B ) Coexpression analysis of 22 PsSPL s in several plant organs. Positive numbers: positive correlations; negative numbers: negative correlations. Red numbers indicate a significant correlation at the 0.05 level
Previous studies demonstrated that some SPL genes are involved in hormone regulation [ 49 , 50 , 51 ]. Therefore, we treated pea plants with five hormones (ABA, JA, SA, GA, and IAA) and evaluated the changes in the transcription levels SPL genes. The expression of most genes was upregulated under ABA, JA, SA, and IAA treatments, whereas most genes were downregulated under GA treatment (Fig. 9 ). PsSPL1 showed the highest expression level under ABA and GA induction, whereas PsSPL13 exhibited the highest expression level under SA and JA induction. PsSPL19 had the highest expression level under IAA induction. Interestingly, these five hormones exhibited peak expression levels at different times. For example, most PsSPLs showed the highest expression levels at 12 h under ABA induction, 1 h under SA induction, and 4 h under JA induction. This finding implies that different hormones regulate the growth and development of pea plants by modulating the expression levels of PsSPLs at different times. We observed upregulation in the expression of PsSPL5 under SA treatment, whereas the expression of this gene was downregulated under other hormone treatments.

Expression analysis of 22 PsSPL s in fruits in response to different hormones (JA, ABA, IAA, SA and GA). ( A ) qRT-PCR was utilized to detect the expression patterns of 22 PsSPLs . Error bars ( n = 3) represent the standard error. Lowercase letters above the bars indicate significant differences ( α = 0.05, LSD) among treatments. ( B ) Coexpression analysis of 22 PsSPL s. Positive numbers = positive correlation; negative numbers = negative correlation. Red numbers indicate a significant correlation at the 0.05 level
Pea is a vegetable crop and an important legume crop [ 1 ]. which is rich in proteins, vitamins and minerals and is widely used in food processing and household consumption [ 2 ]. In agriculture production, peas are commonly used as green manure or feed plants to improve soil quality and can be rotated with other crops to increase agricultural yields [ 52 ]. Additionally, the pea genome is relatively small and the genetic background is relatively simple, so it is easy to study and manipulate [ 53 , 54 ]. Therefore, peas are widely used as a model plant for plant genetics, biology and agricultural research. The SPL gene family plays an important role in fruit development [ 55 , 56 , 57 ]. Investigating the function and regulation mechanism of SPL gene family members during pea fruit development and abiotic stresses can make better understand the molecular mechanisms underlying pea pod formation. Furthermore, it provides a crucial theoretical basis for improving crop varieties and fruit yield and quality.
We identified 22 SPLs in the pea genome and explored the sequence and structural characteristics of these genes. The SPL Protein sizes range from 139 ( PsSPL 3) to 1025 ( PsSPL 9) amino acids. The maximum molecular weight of these proteins was 113.918 kDa (PsSPL18) and the minimum molecular weight was 15.9 kDa (PsSPL4). The number of introns in all PsSPL s ranges from 0 to 14. PsSPL19 has a maximum of 14 introns, while PsSPL5 has no intron structure. Introns play a key role in genome evolution by regulating gene diversity, gene expression regulation, and influencing protein diversity. The intron structure of subfamily II genes is the most complex, suggesting that subfamily II genes may have more functions than other genes. Intron-free genes usually exhibit higher transcription and translation efficiency than intron-containing genes. It is speculated that under stress conditions, the transcriptional regulation process of intron-free genes may be simpler and faster after responding to stimuli [ 58 , 59 , 60 ].
In this study, 22 PsSPL s and their homologous in Arabidopsis thaliana were clustered. The SPLs of both were found to be divided into eight subfamilies, each containing at least one AtSPL gene (Fig. 1 ). Obvious structural and functional differences may be structural and functional differences exists among different SPL subfamilies. There are four fragment repetition events in PsSPL , which may be generated during the evolution of PsSPL family [ 46 , 47 ]. The diversity of sequence length within the PsSPL family may confer adaptability to different environmental and ecological conditions on peas, thus affecting the survival of pea species [ 61 ]. The SPLs clustering results of pea and other six plants showed that PsSPLs were highly homologous to soybean SPLs (Fig. 5 ), although there were obvious differences in morphology and growth habits between the two crops. Phylogenetic analysis shows that subfamily II members are very complex and contain all identified conserved motifs. As a binding protein to gene cis-elements, SPL plays a role as a key regulatory element [ 62 ]. The diversity of motifs may cause PsSPL to bind promoter cis-elements of multiple genes to regulate downstream gene expression in a complex way [ 63 , 64 ], thus exerting diverse functions.
Gene expression analysis is often used for functional prediction to elucidate regulatory mechanisms and biological processes [ 65 ]. qRT-PCR analysis showed that most representative PsSPLs were highly expressed in stems, leaves and flowers, and were involved in the regulation of pod development in pea (Fig. 7 ). Rice OsSPL4 , OsSPL13 and OsSPL16 promote the increase of rice grain width by regulating the level of cytokinin synthesis, and ultimately affect the grain morphological size [ 66 , 67 , 68 ], which seems to support the involvement of SPLs in seed or fruit formation. Under the four non-stress conditions, most of the genes of pea were induced to up-regulate, and the expressions of PsSPL1 , PsSPL5 , PsSPL7 and PsSPL18 of subfamily II were significantly up-regulated in stems under cold and heat stress (Fig. 8 ). It has been reported that AtSPL1 and AtSPL12 , members of II subfamily enhanced the heat resistance of inflorescence by regulating ABA signaling pathway, thereby reducing the sensitivity of flower organs to high temperature stress [ 69 ], suggesting that some SPLs may have similar regulatory mechanisms in the response of pea and Arabidopsis thaliana to heat stress. Moreover, miR156SPL can target AtSPL9 , regulate the binding activity of AtSPL9 with C-REPEAT binding factor 2(CBF2) and induce the expression of AtSPL9 under cold stress, and enhance freezing resistance [ 70 ]. In apples, the miR156/ MdSPL13 module regulates the salt tolerance of apples by targeting the promoter of MdWRKY100 , and overexpression of MdWRKY100 will enhance the salt tolerance of apples [ 71 ], indicating that miR156 usually mediates the response of SPL to salt stress and drought stress. In addition, OsSPL10 imparts drought resistance to rice by regulating the expression of OsNAC2 [ 72 ], while TaSPL6 plays a negative regulatory role in plant drought stress response by reducing the expression of some genes involved in stress response, and overexpression leads to increased sensitivity of wheat to drought stress [ 73 ]. In addition, we predict the cis-acting elements of the PsSPLs promoters in order to find transcription factors that can bind to them, which may be closely related to abiotic stress response (Additional file 6: Table S6 ). All the above studies provide research ideas for revealing the mechanism of PsSPL regulating abiotic stress response. Under four different plant hormone treatments, the expression of most SPLs was increased to varying degrees, mainly at 1 h, 4 h and 12 h after SA, JA and ABA hormone treatments, and the expression of PsSPL1 was increased under ABA and GA induction (Fig. 9 ). Previous studies have shown that in rice, OsSPL12 can directly interact with nine proteins related to GA signal and participate in the regulation of GA synthesis in rice grains [ 49 ], providing clues and references for the mechanism by which PsSPL family participates in GA signal transduction pathway to regulate pod development, promote seed dormancy and inhibit ear germination, etc. PsSPL13 is highly expressed by SA and JA. PsSPL19 may be associated with disease resistance, while IAA induced high expression of PSSPL19 which may promote pod cell elongation or fruit enlargement.
In this study, we identified 22 PsSPLs in the pea genome and conducted a comprehensive analysis of the structure and potential functions of these genes. The 22 PsSPLs were unevenly distributed across seven chromosomes and were classified into eight subfamilies based on homology with AtSPLs . All PsSPLs harbored the SBP domain. Subfamily II exhibited the most complex intron-exon structure and had the highest number of motifs, indicating the functional diversity of genes in this group. The PsSPLs family in peas lacked tandem duplications, but segmental duplications were observed, implying that segmental duplication was involved in the evolution of the gene family. Phylogenetic analysis showed that PsSPLs were highly homologous to soybean SPLs . The expression profile analysis of PsSPLs indicated that these genes may play an important role in the pea growth and development, pod maturation and environmental response. In particular, PsSPL19 is considered as a potential candidate gene for exploring pea breeding.
Data availability
Whole genome sequence information for Pea was obtained from the Ensembl genome website ( http://ensemblgenomes.org ). The seed used in this experiment was Zhongwan 6. The datasets supporting the conclusions of this study are included in the article and in additional files.
Abbreviations
SQUAMOSA promoter-binding protein-like
Pisum sativum L. SPL
quantitative real-time polymerase chain reaction
Arabidopsis thaliana SPL
Hidden Markov Model
isoelectric point
linkage group
days post anthesis
Pandey AK, Rubiales D, Wang Y, Fang P, Sun T, Liu N, Xu P. Omics resources and omics-enabled approaches for achieving high productivity and improved quality in pea (Pisum sativum L.). TAG theoretical and applied genetics Theoretische Und Angewandte Genetik 2021, 134(3):755–76.
Teshome A, Bryngelsson T, Dagne K, Geleta M. Assessment of genetic diversity in Ethiopian field pea (Pisum sativum L.) accessions with newly developed EST-SSR markers. BMC Genet. 2015;16:102.
Article PubMed PubMed Central Google Scholar
Jarambasa T, Regon P, Jyoti SY, Gupta D, Panda SK, Tanti B. Genome-wide identification and expression analysis of the Pisum sativum (L.) APETALA2/ethylene-responsive factor (AP2/ERF) gene family reveals functions in drought and cold stresses. Genetica. 2023;151(3):225–39.
Article CAS PubMed Google Scholar
Bo PT, Bai Y, Dong Y, Shi H, Soe Htet MN, Samoon HA, Zhang R, Tanveer SK, Hai J. Influence of Different Harvesting Stages and Cereals-Legume Mixture on Forage Biomass Yield, Nutritional Compositions, and Quality under Loess Plateau Region. Plants (Basel, Switzerland) 2022, 11(20).
Singh T, Raturi H, Uniyal SP. Effect of Biofertilizer and Mulch on Growth, Yield, Quality and Economics of pea (Pisum sativum L). Indian J Agricultural Res 2021.
Cárcamo HA, Herle CE, Lupwayi NZ. Sitona lineatus (Coleoptera: Curculionidae) larval feeding on Pisum sativum L. affects Soil and Plant Nitrogen. J Insect Sci (Online) 2015, 15(1).
Lahuta LB, Szablińska-Piernik J, Horbowicz M. Changes in metabolic profiles of pea (Pisum sativum L.) as a result of repeated short-term Soil Drought and subsequent re-watering. Int J Mol Sci 2022, 23(3).
Feng K, Hou XL, Xing GM, Liu JX, Duan AQ, Xu ZS, Li MY, Zhuang J, Xiong AS. Advances in AP2/ERF super-family transcription factors in plant. Crit Rev Biotechnol. 2020;40(6):750–76.
Riechmann JL, Heard J, Martin G, Reuber L, Jiang C, Keddie J, Adam L, Pineda O, Ratcliffe OJ, Samaha RR, et al. Arabidopsis transcription factors: genome-wide comparative analysis among eukaryotes. Volume 290. New York, NY): Science; 2000. pp. 2105–10. 5499.
Google Scholar
Yamasaki K, Kigawa T, Inoue M, Tateno M, Yamasaki T, Yabuki T, Aoki M, Seki E, Matsuda T, Nunokawa E, et al. A novel zinc-binding Motif revealed by solution structures of DNA-binding domains of Arabidopsis SBP-family transcription factors. J Mol Biol. 2004;337(1):49–63.
Klein J, Saedler H, Huijser P. A new family of DNA binding proteins includes putative transcriptional regulators of the Antirrhinum majus floral meristem identity gene SQUAMOSA. Mol Gen Genetics: MGG. 1996;250(1):7–16.
CAS PubMed Google Scholar
Shao Y, Zhou HZ, Wu Y, Zhang H, Lin J, Jiang X, He Q, Zhu J, Li Y, Yu H, et al. OsSPL3, an SBP-Domain protein, regulates Crown Root Development in Rice. Plant Cell. 2019;31(6):1257–75.
Article CAS PubMed PubMed Central Google Scholar
He B, Gao S, Lu H, Yan J, Li C, Ma M, Wang X, Chen X, Zhan Y, Zeng F. Genome-wide analysis and molecular dissection of the SPL gene family in Fraxinus mandshurica. BMC Plant Biol. 2022;22(1):451.
Chen X, Zhang Z, Liu D, Zhang K, Li A, Mao L. SQUAMOSA promoter-binding protein-like transcription factors: star players for plant growth and development. J Integr Plant Biol. 2010;52(11):946–51.
Usami T, Horiguchi G, Yano S, Tsukaya H. The more and smaller cells mutants of Arabidopsis thaliana identify novel roles for SQUAMOSA PROMOTER BINDING PROTEIN-LIKE genes in the control of heteroblasty. Development. 2009;136(6):955–64.
Li H, Ma B, Luo Y, Wei W, Yuan J, Zhai C, He N. The Mulberry SPL Gene Family and the response of MnSPL7 to Silkworm Herbivory through activating the transcription of MnTT2L2 in the Catechin Biosynthesis Pathway. Int J Mol Sci 2022, 23(3).
He L, Peng X, Cao H, Yang K, Xiang L, Li R, Zhang F, Liu W. The NtSPL Gene Family in Nicotiana tabacum: genome-wide investigation and expression analysis in response to cadmium stress. Genes 2023, 14(1).
Mao H-D, Yu L-J, Li Z-J, Yan Y, Han R, Liu H, Ma M. Genome-wide analysis of the SPL family transcription factors and their responses to abiotic stresses in maize. Plant Gene. 2016;6:1–12.
Article CAS Google Scholar
Lai D, Fan Y, Xue G, He A, Yang H, He C, Li Y, Ruan J, Yan J, Cheng J. Genome-wide identification and characterization of the SPL gene family and its expression in the various developmental stages and stress conditions in foxtail millet (Setaria italica). BMC Genomics. 2022;23(1):389.
Feng X, Zhou B, Wu X, Wu H, Zhang S, Jiang Y, Wang Y, Zhang Y, Cao M, Guo B, et al. Molecular characterization of SPL gene family during flower morphogenesis and regulation in blueberry. BMC Plant Biol. 2023;23(1):40.
Hou H, Li J, Gao M, Singer SD, Wang H, Mao L, Fei Z, Wang X. Genomic organization, phylogenetic comparison and differential expression of the SBP-box family genes in grape. PLoS ONE. 2013;8(3):e59358.
Liu M, Sun W, Ma Z, Huang L, Wu Q, Tang Z, Bu T, Li C, Chen H. Genome-wide identification of the SPL gene family in Tartary Buckwheat (Fagopyrum tataricum) and expression analysis during fruit development stages. BMC Plant Biol. 2019;19(1):299.
Yang J, Guo Z, Wang W, Cao X, Yang X. Genome-wide characterization of SPL Gene Family in Codonopsis pilosula reveals the functions of CpSPL2 and CpSPL10 in promoting the Accumulation of secondary metabolites and growth of C. Pilosula Hairy Root. Genes 2021, 12(10).
Zhu T, Liu Y, Ma L, Wang X, Zhang D, Han Y, Ding Q, Ma L. Genome-wide identification, phylogeny and expression analysis of the SPL gene family in wheat. BMC Plant Biol. 2020;20(1):420.
Yu N, Yang JC, Yin GT, Li RS, Zou WT. Genome-wide characterization of the SPL gene family involved in the age development of Jatropha curcas. BMC Genomics. 2020;21(1):368.
Ma J, Nie G, Yang Z, Ma S, Fan J, Hu R, Wu F, Zhang X. Genome-wide identification, characterization, and expression profiling analysis of SPL Gene Family during the Inflorescence Development in Trifolium repens. Genes 2022, 13(5).
Wang LY, Yu P, Lyu JY, Hu YF, Han C, Bai MY, Fan M. BZR1 physically interacts with SPL9 to regulate the vegetative phase change and cell elongation in Arabidopsis. Int J Mol Sci. 2021;22(19):10415.
Hu TQ, Manuela D, Xu ML, SQUAMOSA PROMOTER BINDING. PROTEIN-LIKE 9 and 13 repress BLADE-ON-PETIOLE 1 and 2 directly to promote adult leaf morphology in Arabidopsis. J Exp Bot. 2023;74(6):1926–39.
Cui LG, Shan JX, Shi M, Gao JP, Lin HX. The miR156-SPL9-DFR pathway coordinates the relationship between development and abiotic stress tolerance in plants. Plant Journal: Cell Mol Biology. 2014;80(6):1108–17.
Schwarz S, Grande AV, Bujdoso N, Saedler H, Huijser P. The microRNA regulated SBP-box genes SPL9 and SPL15 control shoot maturation in Arabidopsis. Plant Mol Biol. 2008;67(1–2):183–95.
Yao S, Yang Z, Yang R, Huang Y, Guo G, Kong X, Lan Y, Zhou T, Wang H, Wang W, et al. Transcriptional regulation of miR528 by OsSPL9 orchestrates antiviral response in Rice. Mol Plant. 2019;12(8):1114–22.
Jacob A, Lancaster J, Buhler J, Harris B, Chamberlain RD. Mercury BLASTP: accelerating protein sequence alignment. ACM Trans Reconfigurable Technol Syst. 2008;1(2):9.
Marchin M, Kelly PT, Fang J. Tracker: continuous HMMER and BLAST searching. Bioinf (Oxford England). 2005;21(3):388–9.
CAS Google Scholar
Punta M, Coggill PC, Eberhardt RY, Mistry J, Tate J, Boursnell C, Pang N, Forslund K, Ceric G, Clements J, et al. The pfam protein families database. Nucleic Acids Res. 2012;40(Database issue):D290–301.
Letunic I, Bork P. 20 years of the SMART protein domain annotation resource. Nucleic Acids Res. 2018;46(D1):D493–6.
Wang Y, Tang H, Debarry JD, Tan X, Li J, Wang X, Lee TH, Jin H, Marler B, Guo H, et al. MCScanX: a toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012;40(7):e49.
Kumar S, Stecher G, Tamura K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for bigger datasets. Mol Biol Evol. 2016;33(7):1870–4.
Livak KJ. TD Schmittgen 2001 Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods (San Diego Calif) 25 4 402–8.
Alaida MF, Aldhebiani AY. Comparative study of the morphological characteristics of Phoenix dactylifera L. cultivars in Al-Madinah Al-Munawarah-Saudi Arabia. BMC Plant Biol. 2022;22(1):461.
Cenci A. M Rouard 2017 Evolutionary analyses of GRAS transcription factors in Angiosperms. Front Plant Sci 8 273.
Laroche J, Li P, Maggia L, Bousquet J. Molecular evolution of angiosperm mitochondrial introns and exons. Proc Natl Acad Sci USA. 1997;94(11):5722–7.
Oghabian A, Greco D, Frilander MJ. IntEREst: intron-exon retention estimator. BMC Bioinformatics. 2018;19(1):130.
Bailey TL, Boden M, Buske FA, Frith M, Grant CE, Clementi L, Ren J, Li WW, Noble WS. MEME SUITE: tools for motif discovery and searching. Nucleic Acids Res 2009, 37(Web Server issue):W202–208.
Nystrom SL, McKay DJ. Memes: a motif analysis environment in R using tools from the MEME suite. PLoS Comput Biol. 2021;17(9):e1008991.
Yang T, Liu R, Luo Y, Hu S, Wang D, Wang C, Pandey MK, Ge S, Xu Q, Li N, et al. Improved pea reference genome and pan-genome highlight genomic features and evolutionary characteristics. Nat Genet. 2022;54(10):1553–63.
Mascagni F, Usai G, Cavallini A, Porceddu A. Structural characterization and duplication modes of pseudogenes in plants. Sci Rep. 2021;11(1):5292.
Magadum S, Banerjee U, Murugan P, Gangapur D, Ravikesavan R. Gene duplication as a major force in evolution. J Genet. 2013;92(1):155–61.
Article PubMed Google Scholar
Ren Y, Ma R, Fan Y, Zhao B, Cheng P, Fan Y, Wang B. Genome-wide identification and expression analysis of the SPL transcription factor family and its response to abiotic stress in Quinoa (Chenopodium quinoa). BMC Genomics. 2022;23(1):773.
Dong H, Yan S, Jing Y, Yang R, Zhang Y, Zhou Y, Zhu Y, Sun J. MIR156-Targeted SPL9 is phosphorylated by SnRK2s and interacts with ABI5 to enhance ABA responses in Arabidopsis. Front Plant Sci. 2021;12:708573.
Bencivenga S, Simonini S, Benková E, Colombo L. The transcription factors BEL1 and SPL are required for cytokinin and Auxin Signaling during Ovule Development in Arabidopsis. Plant Cell. 2012;24(7):2886–97.
Qin M, Zhang Y, Yang Y, Miao C, Liu S. Seed-specific overexpression of SPL12 and IPA1 improves seed dormancy and grain size in Rice. Front Plant Sci. 2020;11:532771.
Santos CS, Carbas B, Castanho A, Vasconcelos MW, Vaz Patto MC, Domoney C, Brites C. Variation in pea (Pisum sativum L.) seed quality traits defined by Physicochemical Functional properties. Foods (Basel, Switzerland) 2019, 8(11).
Kwon S-J, Brown AF, Hu J, McGee R, Watt C, Kisha T, Timmerman-Vaughan G, Grusak M, McPhee KE, Coyne CJ. Genetic diversity, population structure and genome-wide marker-trait association analysis emphasizing seed nutrients of the USDA pea (Pisum sativum L.) core collection. Genes Genomics. 2012;34(3):305–20.
Kreplak J, Madoui MA, Cápal P, Novák P, Labadie K, Aubert G, Bayer PE, Gali KK, Syme RA, Main D, et al. A reference genome for pea provides insight into legume genome evolution. Nat Genet. 2019;51(9):1411–22.
Zhang X, Dou L, Pang C, Song M, Wei H, Fan S, Wang C, Yu S. Genomic organization, differential expression, and functional analysis of the SPL gene family in Gossypium hirsutum. Mol Genet Genomics. 2015;290(1):115–26.
Zhong H, Kong W, Gong Z, Fang X, Deng X, Liu C, Li Y. Evolutionary analyses reveal diverged patterns of SQUAMOSA promoter binding protein-like (SPL) Gene Family in Oryza Genus. Front Plant Sci. 2019;10:565.
Gandikota M, Birkenbihl RP, Höhmann S, Cardon GH, Saedler H, Huijser P. The miRNA156/157 recognition element in the 3’ UTR of the Arabidopsis SBP box gene SPL3 prevents early flowering by translational inhibition in seedlings. Plant Journal: Cell Mol Biology. 2007;49(4):683–93.
Sánchez D, Ganfornina MD, Gutiérrez G, Marín A. Exon-intron structure and evolution of the lipocalin gene family. Mol Biol Evol. 2003;20(5):775–83.
Xu G, Guo C, Shan H, Kong H. Divergence of duplicate genes in exon–intron structure. 2012, 109(4):1187–92.
Sakharkar M, Long M, Tan TW, de Souza SJ. ExInt: an Exon/Intron database. Nucleic Acids Res. 2000;28(1):191–2.
Carvalho SG, Guerra-Sá R, de Merschmann C. The impact of sequence length and number of sequences on promoter prediction performance. BMC Bioinformatics. 2015;16(19):S5.
Van Roey K, Gibson TJ, Davey NE. Motif switches: decision-making in cell regulation. Curr Opin Struct Biol. 2012;22(3):378–85.
Li J, Mahajan A, Tsai MD. Ankyrin repeat: a unique motif mediating protein-protein interactions. Biochemistry. 2006;45(51):15168–78.
Ciriello GD. Guerra CJBifg, proteomics: a review on models and algorithms for motif discovery in protein-protein interaction networks. 2008, 7 2:147–56.
Ren Y, Ma R, Xie M, Fan Y, Feng L, Chen L, Yang H, Wei X, Wang X, Liu K, et al. Genome-wide identification, phylogenetic and expression pattern analysis of HSF family genes in the Rye (Secale cereale L). BMC Plant Biol. 2023;23(1):441.
Wang S, Wu K, Yuan Q, Liu X, Liu Z, Lin X, Zeng R, Zhu H, Dong G, Qian Q, et al. Control of grain size, shape and quality by OsSPL16 in rice. Nat Genet. 2012;44(8):950–4.
Si L, Chen J, Huang X, Gong H, Luo J, Hou Q, Zhou T, Lu T, Zhu J, Shangguan Y, et al. OsSPL13 controls grain size in cultivated rice. Nat Genet. 2016;48(4):447–56.
Hu J, Huang L, Chen G, Liu H, Zhang Y, Zhang R, Zhang S, Liu J, Hu Q, Hu F, et al. The Elite alleles of OsSPL4 regulate grain size and increase grain yield in Rice. Volume 14. New York, NY: Rice; 2021. p. 90. 1.
Chao LM, Liu YQ, Chen DY, Xue XY, Mao YB, Chen XY. Arabidopsis transcription factors SPL1 and SPL12 Confer Plant Thermotolerance at Reproductive Stage. Mol Plant. 2017;10(5):735–48.
Zhao JL, Shi M, Yu J, Guo CK. SPL9 mediates freezing tolerance by directly regulating the expression of CBF2 in Arabidopsis thaliana . BMC Plant Biol. 2022;22(1):59.
Ma Y, Xue H, Zhang F, Jiang Q, Yang S, Yue PT, Wang F, Zhang YY, Li LG, He P, Zhang ZH. The miR156/SPL module regulates apple salt stress tolerance by activating MdWRKY100 expression. Plant Biotechnol J. 2021;19(2):311–23.
Li YX, Han SC, Sun XM, Ullah KN, Zhong Q, Zhang ZY, Zhang HL, Ming F, Li ZC, Li JJ. Variations in OsSPL10 confer drought tolerance by directly regulating OsNAC2 expression and ROS production in rice. J Integr Plant Biol. 2023;65(4):918–33.
Zhao Y, He JQ, Liu MM, Miao JN, Ma C, Feng YJ, Qian JJ, Li HH, Bi HH, Liu WX. The SPL transcription factor TaSPL6 negatively regulates drought stress response in wheat. Plant Physiol Biochem. 2024;206:108264.
Download references
Acknowledgements
Thanks to all our colleagues for providing useful discussions and technical assistance. Great thanks to the editor and reviewers for critically evaluating the manuscript and providing constructive comments for its improvement.
This research was supported by the Fundamental Research Funds for the Central Universities (2022QNPY54).
Author information
Authors and affiliations.
Minzu University of China, 100010, Beijing, P.R. China
Long Li & Yu ke Geng
School of Food and Biological engineering, Zhengzhou University of Light Industry, 450002, Zhengzhou, P.R. China
Jian bo Xu, Zhi wen Zhu & Xiao zong Wu
College of Agronomy, Hebei Agricultural University, 071001, Baoding, P.R. China
State Key Laboratory of Crop Stress Biology for Arid Areas, College of Plant Protection, Northwest A&F University, 712100, Yangling, Shaanxi, P.R. China
Zhengzhou University of Light Industry, 450002, Zhengzhou, P.R. China
Xiao zong Wu
You can also search for this author in PubMed Google Scholar
Contributions
LL: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Writing-original draft. JBX: Methodology, Software, Investigation, Writing-review & editing. ZWZ: Methodology, Validation. RM: Data curation, Formal analysis, Investigation. XZW: Conceptualization, Supervision. YKG: Funding acquisition, Conceptualization, Supervision. All authors reviewed the manuscript.
Corresponding authors
Correspondence to Xiao zong Wu or Yu ke Geng .
Ethics declarations
Ethics approval and consent to participate.
This article does not contain any studies involving human participants or animals performed by the authors. These methods were carried out in accordance with relevant guidelines and regulations. All experimental protocols were approved by Zhengzhou University of Light Industry.
Consent for publication
Not applicable.
Competing interests
The authors declare that there are no competing interests.
Additional information
Publisher’s note.
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplementary Material 1
Supplementary material 2, supplementary material 3, supplementary material 4, supplementary material 5, supplementary material 6, rights and permissions.
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/ . The Creative Commons Public Domain Dedication waiver ( http://creativecommons.org/publicdomain/zero/1.0/ ) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
Reprints and permissions
About this article
Cite this article.
Li, L., Xu, J.b., Zhu, Z.w. et al. Genome-wide identification and expression analysis of the SPL transcription factor family and its response to abiotic stress in Pisum sativum L. BMC Genomics 25 , 539 (2024). https://doi.org/10.1186/s12864-024-10262-w
Download citation
Received : 15 January 2024
Accepted : 27 March 2024
Published : 31 May 2024
DOI : https://doi.org/10.1186/s12864-024-10262-w
Share this article
Anyone you share the following link with will be able to read this content:
Sorry, a shareable link is not currently available for this article.
Provided by the Springer Nature SharedIt content-sharing initiative
- Pisum sativum. L
- Genome-wide analysis
- Growth and development
- Abiotic stress
- Plant hormone response
BMC Genomics
ISSN: 1471-2164
- Submission enquiries: [email protected]
- General enquiries: [email protected]

An official website of the United States government
The .gov means it’s official. Federal government websites often end in .gov or .mil. Before sharing sensitive information, make sure you’re on a federal government site.
The site is secure. The https:// ensures that you are connecting to the official website and that any information you provide is encrypted and transmitted securely.
- Publications
- Account settings
Preview improvements coming to the PMC website in October 2024. Learn More or Try it out now .
- Advanced Search
- Journal List
- Health Psychol Open
- v.7(2); Jul-Dec 2020
Best practices for stress measurement: How to measure psychological stress in health research
Despite the strong evidence linking psychological stress to disease risk, health researchers often fail to include psychological stress in models of health. One reason for this is the incorrect perception that the construct of psychological stress is too vague and broad to accurately measure. This article describes best practices in stress measurement, detailing which dimensions of stressor exposures and stress responses to capture, and how. We describe when to use psychological versus physiological indicators of stress. It is crucial that researchers across disciplines utilize the latest methods for measuring and describing psychological stress in order to build a cumulative science.
Introduction
Epidemiological studies confirm that both experiencing a greater number of stressful events and reporting high perceived stress over long periods of time are associated with worse mental and physical health, and mortality ( Epel et al., 2018 ). The association between greater stressor exposure and increased disease risk has been replicated with many different types of stressor exposures (e.g. discrimination, caregiving, work stress) and a range of aging-related health outcomes (e.g. cardiovascular disease, metabolic syndrome, mortality). The mechanistic pathways underlying these associations have also been detailed ( Boyce, 2015 ; McEwen, 2015 ; Miller et al., 2009 ). Despite this compelling evidence, however, health researchers often measure stress using unvalidated measures or select a single type of stress to measure, thus either missing entirely or underestimating the role stress plays in predicting disease onset or progression.
One of the main reasons for the lack of sophisticated measurement and inclusion of psychological stress in health models may be the incorrect assumption that stress is too broad and nebulous of a construct to accurately measure. It is true that psychological scientists too often fail to specify what they mean when using the term “stress” or other variants such as “stressor,” “acute stress,” “stress response,” and “stress biomarker.” Social and behavioral scientists tend to use the term loosely, often failing to define it clearly in a manuscript and using it to refer to a range of experiences, from living in poverty to giving a public speech to current negative mood. Kagan (2006) pointed out this lack of specificity, providing a fair critique of the state of the literature. The lack of specificity in language, however, does not represent a true lack of specificity in theoretical or methodological approaches. Although psychological stress researchers have made great strides in differentiating different forms of stress in recent decades, the problem is rather that the language used in journal articles has not always accurately reflected these advancements—and these advancements have been kept within a small, specialized subset of researchers. Thus, the purpose of this article is to provide health researchers across disciplines with a useful update on best practices for measuring stress and offer suggested language for how to describe stress-related constructs with more granular language.
Fundamentals of stress measurement
The term “stress” is an umbrella term representing experiences in which the environmental demands of a situation outweigh the individual’s perceived psychological and physiological ability to cope with it effectively ( Cohen et al., 2016 ). One important distinction in studying stress is to differentiate between exposures to stressful events and the responses to these events. Stressful events or “stressors” are discrete events that can be objectively rated as having the potential to alter or disrupt typical psychological functioning, such as losing your job or getting divorced. Stress responses are the cognitive, emotional, and biological reactions that these stressful events evoke.
Measuring stressor exposures versus stress responses
Stressor exposures can be measured with self-report questionnaires such as a life events checklist, assessed by an interviewer, or objectively determined based on proximity to an event (e.g. living in NYC during the September 11 terrorist attacks). The Life Events and Difficulties Schedule (LEDS; Brown and Harris, 1978 ) is a structured interview protocol that is considered the gold standard for assessing stressor exposure across someone’s lifetime. This interview protocol is time intensive in both the data collection and data processing stages. To streamline the process of capturing stressor exposures across the life span, a computer-assisted methodology was developed (e.g. The Stress and Adversity Inventory [STRAIN]; Slavich and Shields, 2018 ). In both the LEDS and the STRAIN, participants are asked whether they have experienced a range of stressful life events at any point in their life. For each endorsed stressor, they are asked follow-up questions to provide greater context about the experience (e.g. how old were you when it happened, how long did it go on for, how stressful or threatening was it). The LEDS requires a trained interviewer to administer the measure, while the STRAIN can be completed either by an interviewer or by participants themselves. The LEDS also relies on blind raters to score the severity of a stressor using this contextual information, while the STRAIN relies on the participants reporting of event severity. The STRAIN’s automated structure of follow-up questions allows the respondent to complete the interview much more quickly than the LEDS and reduces data processing time. Both measures provide a comprehensive assessment of stressor exposures across the lifespan, and use different methods to determine the severity of these experiences.
An individual’s response to the stressor sometimes matters more than mere exposure to it, particularly when it comes to the impact of the stressor on physical health. For example, caregiving for a family member with a debilitating illness is often considered a chronic stressor because of the constant physical and emotional demands. There is a significant amount of research examining the impact of being a dementia caregiver, in particular, given the large increase in the number of family dementia caregivers as the population ages in the United States. In fact, the Alzheimer’s Association estimated in 2018 that there were over 16 million family caregivers providing an estimated 18.5 billion hours of care to people with Alzheimer’s or other dementias ( Alzheimer’s Association, 2019 ). Empirical evidence has shown that family caregivers of Alzheimer’s patients have worse physical and mental health compared to age-matched non-caregivers ( Kiecolt-Glaser et al., 1987 ; Vitaliano et al., 2003 ). However, not every caregiver’s health is damaged by their caregiving role ( Roth et al., 2015 ). This may be because the negative impact of caregiving is caused by individuals’ subjective response to the caregiving situation, not from the mere exposure of being a caregiver. Thus, a better predictor of health decline would be the degree to which caregivers report high levels of psychological burden from their caregiving role. Empirical evidence supports this perspective; for example, Alzheimer’s caregivers who reported emotional distress or physical strain from caregiving had 63 percent greater mortality than caregivers who reported no distress ( Schulz et al., 1999 ).
Stress responses can be measured with self-report measures, behavioral coding, or via physiological measurements. These responses include emotions, cognitions, behaviors, and physiological responses instigated by the stressful stimuli. One of the simplest ways to measure stress responses is through self-reports of perceived stress related to a specific stressor or to one’s life circumstances ( Cohen et al., 1983 ). For example, the Perceived Stress Scale is a 10-item self-report measure that captures an individual’s perception of how overwhelmed they are by their current life circumstances. Responses to acute stressors have traditionally been studied in controlled laboratory settings in order to capture responses that unfold within minutes of stressor exposure (e.g. emotional and physiological reactivity to an acute stress task). A commonly used acute stress paradigm is the Trier Social Stress Test (TSST), a standardized laboratory stress task in which participants give a speech and perform mental arithmetic in front of judges ( Kirschbaum et al., 1993 ). The TSST reliably evokes an acute stress response for the majority of participants. Outside of the laboratory, new technology has enhanced our ability to capture real-time stress responses in daily life using mobile phones and wearables, which many researchers are now doing. Considering the impact of both stressor exposure and stress responses on health may improve the prediction of health outcomes, as many models of stress propose that the stress response mediates the effect of stress exposures on health outcomes ( McEwen, 1998 ; Wheaton et al., 2013 ).
Selecting stress measures
Due to constraints on participant burden and other considerations, difficult choices about which type of stress to measure need to be made by researchers. Common types of psychological stress measured using self-report questionnaires in adult samples are major life events, traumatic events, early life stress exposure, and current chronic or perceived stress in various domains (i.e. loneliness, marital discord, experiences of discrimination, work stress, financial strain, neighborhood safety and cohesion, and current perceived stress). The choice of which type of stressor exposure to measure depends on what is most relevant to the study population, the specific research question, and the hypothesized mechanisms linking that stress type to the outcome of interest. To begin the selection, consider first what is the most relevant stress type(s), given the sample’s demographic makeup. For example, measures that capture religious persecution or combat exposure would be particularly important for a sample living in a conflict zone, while the amount of overwhelm related to being a parent (parenting stress) may be most relevant for a sample of mothers caring for their child who has an autism spectrum disorder. In both cases, it would also be important to measure types of stressors that may not be directly related to the circumstances—such as levels of loneliness and financial strain. Capturing a range of stressor types reduces the likelihood that the individual’s psychological and social distress is underestimated.
Stressor and stress response characteristics
In addition to identifying stressor type(s) of interest, there are several key measurement considerations when choosing specific measures of stress to include in studies or analyzing existing stress measure data. These considerations include characteristics of the stressor or response (e.g. timescale, types of stressor response) as well as measurement characteristics (e.g. life stage of exposure and measurement assessment window). We briefly describe these aspects below (see Epel et al., 2018 for further discussion).
Timescale of the stressor
Stressors generally take place along the following timescales: chronic stressors, life events, daily events/hassles, and acute stress. Table 1 provides definitions for each of these timescales. It is important to note that naturalistic experiences of stress rarely fall neatly into one category. For example, death of a loved one is often considered a major life event but, depending on the cause of death, may also be considered a chronic stressor, such as if the family member was sick for years or months before the death. Similarly, arguments with a spouse may be considered an acute stressor, but if they happen every day they may be considered chronic. There is a significant amount of gray area between categories. Researchers should first make a thoughtful attempt to pick the category that best aligns with the stressor and with the way that stressor type has been described in past research, and then describe the exposure with as much specificity as possible.
Types of stress by timescale.
Types of stress response
Responses to stressor exposures provide additional useful information beyond measuring stressor exposure alone. Stress responses include psychological, behavioral, cognitive, and physiological reactions related to the stressor exposure that can occur before, during, or after the exposure. Psychological stress responses include specific emotions triggered by the stressor, as well as efforts to regulate that emotion ( Gross, 2002 ). Behavioral responses include coping behaviors such as smoking or seeking social support. Cognitive responses include appraisals of the exposure (e.g. as a threat versus challenge; Blascovich and Mendes, 2010 ) and perseverative cognitions (e.g. rumination Brosschot et al., 2005 ). Physiological responses include immune, autonomic, neuroendocrine, and neural changes related to stressor exposure. Further details about the various stress responses deserve more attention than can be described here ( Epel et al., 2018 ). As a part of selecting stress measures, researchers should identify the type of stress response that is most relevant for their research question and sample. Often, studies will assess multiple types of stress responses simultaneously.
Additional characteristics of the stressor
There are additional stressor exposure attributes that can be described and captured to thoroughly assess the exposure. These include, but are not limited to, duration, severity, controllability, life domain, the target of the stressor (e.g. self, close other), and the potential of the stressor to elicit specific harmful emotional responses (e.g. social status threat). Lack of control, social status threat, and stressor severity have been identified as potent attributes that predict worse outcomes across a range of stressor types and scenarios.
Characteristics of stress measurement
Life stage during stressor exposure.
In addition to the timescale of the stressor, another important characteristic of stressor exposure is the developmental or life stage during which the stressor occurs. Knowing the person’s age during the exposure informs hypotheses about which psychological and biological processes the stressor may have impacted. This is because developing systems are more open to environmental cues and are thus more likely to be impacted by stress exposure. “Sensitive periods” are specific time points in the life course during which physiological systems are maximally influenced by external environmental factors, and thus stressor exposure can have a particularly strong influence on development ( Knudsen, 2004 ). Sensitive periods during which stress may have the greatest effect are likely: prenatal ( Van Den Bergh et al., 2005 ; Weinstock, 2001 ), before age 5 ( Zeanah et al., 2011 ), during puberty ( Fuhrmann et al., 2015 ), entry into parenthood ( Saxbe et al., 2018 ), and during menopause ( Gordon et al., 2015 ). Identifying and measuring stress during sensitive periods could greatly increase our understanding of who is at risk for the negative effects of stress, the mechanistic pathways linking stress exposure to health decline, and where and how to focus intervention efforts.
Measurement assessment window
The window of measurement is also essential to consider to avoid measurement error and improve specificity in hypotheses. Measures can ask about stressors and stress responses across a wide range of time frames, such as in the present moment, over the course of that day, the past week, the past month, the past year, in childhood, or across the entire lifespan. For example, there are fundamental differences in a measure that ask participants to report on stress exposure in the past month versus across their lifespan. The latency between stressor exposure and measurement is crucial, as retrospective autobiographical reports are prone to bias and error, especially when there have been years or decades since the exposure in question ( Bradburn et al., 1987 ; Hardt and Rutter, 2004 ). In addition to the latency between exposure and measurement, several other factors can impact the accuracy of retrospective reports, such as mental state at the time of recall and the emotional salience of a given memory ( Shiffman et al., 2008 ). This may lead to overestimating the frequency of emotionally salient stressors and underestimating the frequency of more mundane, daily stressors ( Bradburn et al., 1987 ; Shiffman et al., 2008 ). For these reasons, it can be beneficial to measure stressor exposure and responses in close proximity to their occurrence whenever possible.
The experimental studies examining acute stressor exposure and responses, there are additional considerations with the measurement assessment window. Because the timing of stressor exposure is controlled, researchers can begin measuring psychological, behavioral, and physiological states prior to the stressor exposure and continue measuring throughout and after exposure. By measuring responses before, during, and after exposure, researchers can access (and predict) anticipation of and recovery from the stressor exposure.
Summary of steps for selecting stress measures
There are of course numerous considerations for selecting the appropriate stress measure for your study. In sum, researchers should identify the type or types of stress that are most relevant to their research question and sample. Stress measure selection should then be refined based on characteristics of the stressor and/or stress response that the researcher intends to measure, such as the timescale, the type(s) of stress responses the researcher is interested in, and other attributes of the stressor (e.g. duration, severity, controllability). Selection of stress measures should also account for measurement characteristics, such as the life stage during stressor exposure and the measurement assessment window (e.g. framing of questions, timing of assessment relative to occurrence of the stressor).
Beyond these stress-specific considerations, researchers should also follow general best practices for measure selection. For example, validated scales should be used when available. The Stress Measurement Network Toolbox provides a resource for validated measures of different types of stress that has beeen curated by experts ( https://stressmeasurement.org ). Measures should also be selected based on the uniqueness of the sample, and hile validated scales are preferred, some samples or exposures may require researchers to develop a new scale or modify an existing scale to fit their needs. These practical steps for selecting a stress measure are summarized in Table 2 .
Summary of steps for choosing appropriate stress measures.
Compelling evidence linking stress to physical health
The types of stress that have the most consistent and compelling relationships with disease risk and mortality are acute stress reactivity, early life stress, work or occupational stress, and social isolation/loneliness. A comprehensive review of these literatures is outside the scope of the present article; however, the following section highlights foundational studies linking these stress types physical health, with a particular emphasis on cardiovascular disease (because it is the leading cause of death in developed countries) and mortality. Effect sizes are included where possible, as are references to reviews and meta-analyses for further reading. Of note, we do not review the literature here on the impact of cumulative life stress (aggregate number of stressor exposures and/or intensity of stress responses over one’s life course). Despite initial compelling work on the impact of cumulative life stress on cardiovascular disease outcomes, this area of research is still in its infancy, with a need for measurement approaches to be unified across research studies to allow for building of a collective science ( Albert et al., 2013 ; Slopen et al., 2018 ).
Research on acute stress reactivity and physical health
Decades of research have shown that heightened cardiovascular reactivity and delayed recovery to acute stressors are prospectively associated with increased cardiovascular disease risk ( Brosschot et al., 2005 ; Chida and Steptoe, 2010 ; Steptoe and Marmot, 2005 ). One of the earliest studies in this area was a longitudinal study of healthy adult men (age 45–55; n = 279) in which those classified as “hyper-reactors” (defined as > 20 mmHg increase in diastolic blood pressure to the cold pressor acute stress task) were 2.4 times more likely to have a myocardial infarction or die from cardiovascular disease in the following 20 years than men who showed a rise of < 20 mmHg ( Keys et al., 1971 ). Cortisol and inflammatory responses to acute stressors have also been shown to prospectively predict incident hypertension ( Hamer and Steptoe, 2012 ; Steptoe et al., 2016 ). Heightened reactions and prolonged recovery time periods may be driven by perseverative cognitions before (worrying) and after (rumination) stressor exposure ( Brosschot et al., 2005 , 2006 ). Despite the evidence linking reactivity to disease outcomes, the clinical meaningfulness of these associations is still debated ( Treiber et al., 2003 ). Importantly, a blunted response to an acutely stressful situation (sometimes termed a “hyporeactive response”), is also linked to worse health ( Carroll et al., 2017 ). For example, in a sample of 725 healthy adults from the Dutch Famine Birth Cohort Study, decreased cardiovascular and/or cortisol response to the acute stressor was associated with obesity, risk of becoming obese, depressive symptoms, anxiety, and poor self-rated and functional health ( De Rooij, 2013 ). In addition, there are several other reactivity patterns that have been hypothesized to represent maladaptive response profiles such as lack of habituation when exposed to repeated stressors of the same kind (see McEwen, 1998 ). Thus, the clinical meaningfulness of different stress reactivity profiles is largely debated.
Research on early life stress and physical health
The evidence linking early life stress to increased adult disease risk and mortality is strong. A foundational study in this area, the Adverse Childhood Experiences (ACE) Study, included nearly 10,000 adults and demonstrated that a greater number of self-reported retrospective adverse childhood experiences (e.g. physical abuse, living with an alcohol-dependent adult, witnessing violence) was positively associated in a graded relationship with the presence of ischemic heart disease, cancer, chronic lung disease, skeletal fractures, and liver disease, after controlling for demographic factors ( Felitti et al., 1998 ). Convincingly, reporting seven or more ACE was associated with three times the likelihood of heart disease compared to reporting no ACE ( Dong et al., 2004 ). These findings have been so compelling that significant changes in clinical and educational settings have been undertaken in recent years to recognize the role that early trauma has on current and future cognitive, socio-emotional, and behavioral outcomes for both children and adults.
Research on work stress and physical health
Epidemiological studies consistently demonstrate associations between high work stress and worse physical and mental health. One of the most widely studied models of work stress is job strain, which is a combination of high demands (workload and intensity) and low control ( Karasek, 1979 ). Decades of research has linked high job strain to anxiety and depression, increased blood pressure (BP), cardiovascular events, and metabolic syndrome ( Chandola et al., 2006 ; Landsbergis et al., 2013 ; Madsen et al., 2017 ; Nyberg et al., 2013 ). An analysis of the Whitehall II study cohort found that chronic work stress was associated with coronary heart disease (CHD) risk, with the associations strongest in participants under 50 (RR = 1.68, 95% CI 1.17–2.42). Other components of work stress, such as effort-reward imbalance, also predict cardiovascular disease risk ( Dragano et al., 2017 ).
Research on social isolation, loneliness, and physical health
A meta-analysis of decades of work on social isolation and loneliness found that being socially isolated, lonely, and/or living alone corresponded to an average of 29 percent, 26 percent, and 32 percent increased likelihood of mortality ( Holt-Lunstad et al., 2015 ). The mortality risk for the most socially isolated adults in the National Health and Nutritional Examination Survey (hazard ratio (HZ) = 1.62 for men, HZ = 1.75) was found to be comparable to the risk of smoking (HR = 1.72 for men, HZ = 1.86) and having high BP (HR = 1.16 for men, HR = 1.32 for women) ( Pantell et al., 2013 ). These strong relationships suggest that meaningful connection with others is an essential component of health and well-being. Several short measures have been created to capture this important social determinant of health, including a validated three-item measure of loneliness ( Hughes et al., 2004 ).
Biological pathways from stress to disease
There are numerous plausible biological pathways linking stress to cardiovascular disease, with most of the current evidence pointing to stress-related alterations in the immune, autonomic, and neuroendocrine systems. The brain networks that orchestrate stress-induced changes in these peripheral systems have also been identified ( Gianaros and Wager, 2015 ; Gianaros and Jennings, 2018 ), and can be described as the systems related to threat processing, safety processing, and social cognition ( Muscatell and Eisenberger, 2012 ). One widely accepted stress-disease model is the “wear and tear” hypothesis ( Charles et al., 2013 ; McEwen, 1998 ; Selye, 1956 ). This hypothesis is centered on the postulation that prolonged or repeated stress prematurely depletes the finite amount of “adaptational energy” of the organism, decreasing the body’s ability to successfully adapt to environmental challenges ( Selye, 1956 ). In this model, stressful events cause stress responses that involve activation of physiologic systems. In the short term, mobilizing physiological resources to respond to a discrete event or threat is adaptive. In the long term, however, frequent and repeated mobilization of these resources wears down these response systems and maladaptive patterns appear ( McEwen, 1998 ). The “wear and tear” hypothesis is theoretically compelling, but currently lacks definitive empirical support. This is because we do not currently have data that demonstrates the slow degradation of multiple physiological systems over decades in humans, an effort that requires tremendous investment. Instead, most studies have chosen one or maybe two physiological systems to measure to try to capture degradation or maladaptive responses to stressors, thus providing support, but not direct evidence for the “wear and tear” hypothesis. Other potential pathways include stress-related changes in endothelial function, elevated chronic inflammation, metabolic dysfunction, changes in DNA repair, changes in gene expression, and telomere shortening. These are all exciting areas of research, some of which fit in to the “wear and tear” hypothesis (e.g. telomere shortening; Epel et al., 2004 ) and others that suggest alternate processes (e.g. biological embedding of early experiences; Miller et al., 2011 ). These pathways are relevant for numerous chronic diseases beyond cardiovascular disease.
Associations between stress and immune system functioning are especially relevant given that the major diseases of aging in the United States are mediated, in part, through the immune system. The top three leading causes of death in the United States—cardiovascular disease, cancer, and chronic lower respiratory disease—all share the common thread of being characterized by elevated chronic inflammation ( Aghasafari et al., 2019 ; Golia et al., 2014 ; Grivennikov et al., 2010 ). Because of this common thread, chronic systemic inflammation has become a recent focus of health research. Stress exposure has been examined extensively as a predictor of increased systemic inflammation. Indeed, elevated systemic inflammation has been found in those experiencing chronic stress like caregivers ( Gouin et al., 2008 ), immediately after a stressful life event like death of a loved one ( Cohen et al., 2015 ), historical stress like childhood adversity ( Slopen et al., 2010 , 2012 ), daily stress ( Chiang et al., 2012 ), and in response to lab-based stress tasks ( Marsland et al., 2017 ). A short-term inflammatory response to stress is thought to be adaptive because it involves recruiting immune cells to the site of a real or potential injury in order to heal wounds resulting from stressor exposure. However, when there is no wound to heal, as is the case with psychosocial stressor exposure, repeated or exaggerated inflammatory responses may cause long-term damage and contribute to disease processes ( Black and Garbutt, 2002 ; Miller et al., 2002 ; Rohleder, 2014 ).
Is there an “objective” way to measure stress?
Stress and health researchers have searched for many years for a single biological indicator that someone is “under stress.” However, there is no single stress-specific biomarker. This is likely because acute stress is not the only state that evokes reliable biological changes (e.g. increased heart rate and BP). Other non-acute stress states, such as feeling excited, focusing attention on non-negative affect inducing stimuli, or exercising, also trigger biological responses that are similar to those evoked by negative affect inducing acute stressors like increased heart rate and blood pressure. This is even true for what is often termed the “stress hormone,” cortisol—not all cortisol increases are triggered by increases in psychological stress responses, nor does every experience that people perceive as “stressful” cause cortisol to rise ( Dickerson and Kemeny, 2004 ).
While measuring stress-related biomarkers may not provide a perfect indicator of whether someone is under stress or not, there are still compelling reasons to include these biomarkers in research studies of stress and health. Stress-related biomarkers are objectively measured biological indicators of physiological processes that are either implicated in the pathway from stress to disease or serve as a marker of that process. In typical models of the stress-health relationship, the stressful event (X) leads to a biological change (Y) that then leads to the disease state or related outcome (Z). Stress-related biomarkers can be the variable inserted in any component (X, Y, or Z) of this model; examples of the stress-related biomarker in each part of this basic model are shown in Figure 1 . In example A, the biomarker serves as a mediator, or a part of the causal pathway between a stressor and a health outcome. In example B, the biomarker serves as a predictor of stress-related psychosocial and behavioral processes that ultimately impact health outcomes. In example C, the biomarker serves as an outcome of psychological and physiological responses to a traumatic stressor. The way a biomarker is conceptualized (e.g. as a mediator, predictor, or outcome) depends on the research question and study methods. As such, choosing a stress-related biomarker to include in a study depends on the design of the study and the outcomes of interest. Table 3 provides further tools for how to choose the appropriate biomarker. It is also important to keep in mind that a biomarker may not be needed to answer a research question, despite the desire for a seemingly “objective” indicator of stress or stress reduction.

Examples of how stress-related biomarkers can be modeled as either the predictor, the mediator, or the outcome in research studies.
Essential questions for following best practices in choosing an appropriate stress-related biomarker.
One area of research that requires particularly careful consideration of biomarker selection is when biomarkers are used as an outcome in psychosocial intervention trials. The scientific community is often eager to find an objective biological indicator that a psychosocial intervention can improve health; this is typically done by measuring improvement in a biomarker from pre- to post-intervention. There has been a trend in recent years toward using changes in biomarkers as an indicator of an intervention’s success, rather than relying on subjective psychological reports of well-being. This approach is problematic for several reasons, including variability in baseline biomarker profiles, unknown reliability of biomarker assessment over time, unknown stability of these changes, and lack of evidence for the long-term impact of small changes in stress-related biomarkers on disease risk. Therefore, biomarkers should not replace self-report, behavioral, and cognitive outcomes as primary outcomes in psychosocial intervention trials aimed at reducing stress or related goals.
Variability in exposures and responses
Despite stress exposure being an inevitable part of life, not everyone develops stress-related illnesses at the same speed. One primary reason for this is that stress exposures are not distributed evenly across social groups. Women, young adults, members of racial-ethnic minority groups, divorced and widowed persons, and poor and working-class individuals report greater chronic stress and cumulative stress exposure across their lives ( Thoits, 2010 ). In addition, recent research has demonstrated that both psychological and physiological stress responses vary remarkably within and between people. While the physiological systems that are activated in response to a stressor are generally universal and non-specific as initially proposed by one of the founders of the field of stress, Hans Selye (1956) , the pattern of these responses vary considerably in terms of the degree of the system’s activation and how long the systems are activated for. Individual-level differences and environmental contexts interact to influence the psychological and physiological stress response trajectories. These include socioeconomic and cultural factors, genetic and developmental factors, historical and current stressors, stable protective factors, and health behaviors. A model integrating these different levels of experience is presented by our group in detail in Epel et al. (2018) and reprinted here with permission ( Figure 2 ).

Transdisciplinary model of psychological stress: Integrating contextual, historical, habitual, and acute stress processes.
Figure 2 presents a transdisciplinary model that describes psychological stress as encompassing as a set of interrelated processes. The figure illustrates that stressors are experienced within the context of a person’s life, represented by the contextual factors in the blue triangle. These contextual factors include individual-level characteristics such as personality and demographics, the environment in which one lives, current and past stressor exposures, and protective factors—all of which combine to determine the baseline allostatic state of physiological regulation, and the lens through which stressors are perceived and assigned meaning. Contextual factors and habitual processes together influence psychological and physiological responses to acute and daily stressors. These responses, if dysregulated, are thought to lead to allostatic load and ultimately biological aging and early disease. Reprinted from Frontiers in Neuroendocrinology ( Epel et al., 2018 ).
Advanced statistical models can be used to examine variability in stress responses (both psychological and physiological) within and between people ( Bryk and Raudenbush, 1987 ; McArdle and Epstein, 1987 ). Within-person variability in stress responses means that a person’s response to a stressor within one life domain (e.g. work) does not necessarily predict how they will respond to a stressor within another life domain (e.g. family). Between-person variability means that different people respond to the same stressor in a variety of ways. As an example of variability in psychological stress responses, in a sample of 1,532 healthy adults from the Changing Lives of Older Couples prospective study, psychological responses to the death of one’s spouse took on four discrete trajectories (e.g. chronic grief, chronic depression, temporary depression, resilient), suggesting that there is not one universal pattern for spousal grief ( Galatzer-Levy and Bonanno, 2012 ). Cortisol can be used as an example of variability in physiological stress responses Cortisol generally increases in response to laboratory-based acute stress tasks if they are uncontrollable and characterized by social-evaluative threat ( Dickerson and Kemeny, 2004 ), such as the TSST described earlier ( Kirschbaum et al., 1993 ). However, around 30 percent of people do not mount a cortisol response, and there is tremendous variability in the size of the response. Individual-level predictors of this variability include age, gender, sex steroid levels, smoking, coffee, and alcohol consumption ( Kudielka et al., 2009 ). Interestingly, these differences are not driven by differences in the emotional responses to the task as acute stressors are not strongly correlated to the physiological responses. In a review of 49 acute stress studies, only 25 percent reported a significant correlation between the two emotional and physiological responses ( Campbell and Ehlert, 2012 ).
Empirical evidence supports a strong relationship between psychological stress and disease development. These studies may be underestimating the impact of stressor exposure and the stress response on health, given that measuring these constructs has been challenging and limited. Recent work in the stress field has identified important aspects of psychological stress to capture in order to fully test the role that psychological stress plays in predicting disease; these include capturing the specific type(s) of stressor exposure, a wide range of psychological, cognitive, behavioral, and physiological responses to the exposure, and contextual and individual-level factors that moderate the impact of the exposure and response. In this article, we identified ways for researchers to improve the language specificity when describing stress measures and offered guidance on how to choose the appropriate stress measure. We encourage the adoption of more precise language when writing about stress in academic papers, more careful selection of stress measures, with a focus on validated measures when possible, and theoretically driven integration of mechanistic pathways linking stress to health outcomes. The ultimate goal of having sophisticated research on the relationship between stress, health, and well-being is to develop evidence-based ways to help people thrive in our stress-filled world.
Acknowledgments
Members of the Stress Measurement Network provided essential input on the thoughts presented here, and we thank them for their contribution.
Conflict of Interests: The author(s) declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding: The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was funded by the National Institute on Aging of the National Institutes of Health [R24AG048024; K01AG057859].

- Aghasafari P, George U, Pidaparti R. (2019) A review of inflammatory mechanism in airway diseases . Inflammation Research 68 : 59–74. [ PubMed ] [ Google Scholar ]
- Albert MA, Slopen N, Williams DR. (2013) Cumulative psychological stress and cardiovascular disease risk: A focused review with consideration of black-white disparities . Current Cardiovascular Risk Reports 7 : 318–325. [ Google Scholar ]
- Almeida DM. (2005) Resilience and vunerability to daily stressors assessed via diary methods . Current Directions in Psychological Science 14 ( 2 ): 64–68. [ Google Scholar ]
- Alzheimer’s Association (2019) 2019 Alzheimer’s disease facts and figures . Alzheimer’s & Dementia 15 ( 3 ): 321–387. [ PubMed ] [ Google Scholar ]
- Black PH, Garbutt LD. (2002) Stress, inflammation and cardiovascular disease . Journal of Psychosomatic Research 52 : 1–23. [ PubMed ] [ Google Scholar ]
- Blascovich JJ, Mendes WB. (2010) Social psychophysiology and embodiment . In: Fiske ST, Gilbert DT, Lindzey G. (eds) Handbook of Social Psychology . Hoboken, NJ: John Wiley & Sons, pp. 194–227. [ Google Scholar ]
- Boyce WT. (2015) Epigenomics and the unheralded convergence of the biological and social sciences . In: Kaplan R, Spittel M, David DH. (eds) Population Health: Behavioral and Social Science Insights . Rockville, MD: Agency for Healthcare Research and Quality and Office of Behavioral and Social Sciences Research, National Institutes of Health, pp. 219–232. [ Google Scholar ]
- Bradburn N, Rips L, Shevell S. (1987) Answering autobiographical questions: The impact of memory and inference on surveys . Science 236 ( 4798 ): 157–161. [ PubMed ] [ Google Scholar ]
- Brosschot JF, Gerin W, Thayer JF. (2006) The perseverative cognition hypothesis: A review of worry, prolonged stress-related physiological activation, and health . Journal of Psychosomatic Research 60 : 113–124. [ PubMed ] [ Google Scholar ]
- Brosschot JF, Pieper S, Thayer JF. (2005) Expanding stress theory: Prolonged activation and perseverative cognition . Psychoneuroendocrinology 30 ( 10 ): 1043–1049. [ PubMed ] [ Google Scholar ]
- Brown G, Harris T. (1978) Social Origins of Depression: A Study of Psychiatric Disorders in Women . New York: The Free Press. [ Google Scholar ]
- Bryk AS, Raudenbush SW. (1987) Application of Hierarchical Linear Models to Assessing Change . Psychological Bulletin 101 : 147–158. [ Google Scholar ]
- Campbell J, Ehlert U. (2012) Acute psychosocial stress: Does the emotional stress response correspond with physiological responses? Psychoneuroendocrinology 37 ( 8 ): 1111–1134. [ PubMed ] [ Google Scholar ]
- Carroll D, Ginty AT, Whittaker AC, et al. (2017) The behavioural, cognitive, and neural corollaries of blunted cardiovascular and cortisol reactions to acute psychological stress . Neuroscience and Biobehavioral Reviews 77 : 74–86. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Chandola T, Brunner E, Marmot M. (2006) Chronic stress at work and the metabolic syndrome: Prospective study . BMJ: British Medical Journal 332 : 521–525. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Charles ST, Piazza JR, Mogle J, et al. (2013) The wear and tear of daily stressors on mental health . Psychological Science 24 ( 5 ): 733–741. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Chiang JJ, Eisenberger NI, Seeman TE, et al. (2012) Negative and competitive social interactions are related to heightened proinflammatory cytokine activity . Proceedings of the National Academy of Sciences of the United States of America 109 ( 6 ): 1878–1882. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Chiang JJ, Turiano NA, Mroczek DK, et al. (2018) Affective reactivity to daily stress and 20-year mortality risk in adults with chronic illness: Findings from the National Study of daily experiences . Health Psychology 37 ( 2 ): 170–178. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Chida Y, Steptoe A. (2010) Greater cardiovascular responses to laboratory mental stress are associated with poor subsequent cardiovascular risk status: A meta-analysis of prospective evidence . Hypertension 55 ( 4 ): 1026–1032. [ PubMed ] [ Google Scholar ]
- Chida Y, Hamer M, Wardle J, et al. (2008) Do stress-related psychosocial factors contribute to cancer incidence and survival? Nature Clinical Practice Oncology 5 ( 8 ): 466–475. [ PubMed ] [ Google Scholar ]
- Cohen M, Granger S, Fuller-Thomson E. (2015) The association between bereavement and biomarkers of inflammation . Behavioral Medicine 41 ( 2 ): 49–59. [ PubMed ] [ Google Scholar ]
- Cohen S, Gianaros P, Manuck S. (2016) A stage model of stress and disease . Perspectives on Psychological Science 11 ( 4 ): 456–463. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Cohen S, Janicki-Deverts D, Miller GE. (2007) Psychological stress and disease . Journal of the American Medical Association 298 : 1685–1687. [ PubMed ] [ Google Scholar ]
- Cohen S, Kamarck T, Mermelstein R. (1983) A global measure of perceived stress . Journal of Health and Social Behavior 24 ( 4 ): 385–396. [ PubMed ] [ Google Scholar ]
- De Rooij SR. (2013) Blunted cardiovascular and cortisol reactivity to acute psychological stress: A summary of results from the Dutch Famine Birth Cohort Study . International Journal of Psychophysiology 90 ( 1 ): 21–27. [ PubMed ] [ Google Scholar ]
- Dickerson SS, Kemeny ME. (2004) Acute stressors and cortisol responses: A theoretical integration and synthesis of laboratory research . Psychological Bulletin 130 ( 3 ): 355–391. [ PubMed ] [ Google Scholar ]
- Dong M, Giles WH, Felitti VJ, et al. (2004) Insights into causal pathways for ischemic heart disease: Adverse childhood experiences study . Circulation 110 : 1761–1766. [ PubMed ] [ Google Scholar ]
- Dragano N, Siegrist J, Nyberg ST, et al. (2017) Effort-reward imbalance at work and incident coronary heart disease: A multicohort study of 90,164 individuals . Epidemiology 28 ( 4 ): 619–626. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Epel E, Blackburn EH, Lin J, et al. (2004) Accelerated telomere shortening in response to life stress . Proceedings of the National Academy of Sciences of the United States of America 101 ( 49 ): 17312–17315. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Epel E, Crosswell A, Mayer S, et al. (2018) More than a feeling: A unified view of stress measurement for population science . Frontiers in Neuroendocrinology 49 : 146–169. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Felitti VJ, Anda RF, Nordenberg D, et al. (1998) Household dysfunction to many of the leading causes of death in adults the Adverse Childhood Experiences (ACE) Study . American Journal of Preventive Medicine 14 ( 4 ): 245–258. [ PubMed ] [ Google Scholar ]
- Fuhrmann D, Knoll LJ, Blakemore SJ. (2015) Adolescence as a Sensitive Period of Brain Development . Trends in Cognitive Sciences 19 : 558–566. [ PubMed ] [ Google Scholar ]
- Galatzer-Levy IR, Bonanno GA. (2012) Beyond normality in the study of bereavement: Heterogeneity in depression outcomes following loss in older adults . Social Science & Medicine 74 ( 12 ): 1987–1994. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Gawronski KAB, Kim ES, Miller LE. (2014) Potentially traumatic events and serious life stressors are prospectively associated with frequency of doctor visits and overnight hospital visits . Journal of Psychosomatic Research 77 ( 2 ): 90–96. [ PubMed ] [ Google Scholar ]
- Gianaros P, Wager T. (2015) Brain-body pathways linking psychological stress and physical health . Current Directions in Psychological Science 24 ( 4 ): 313–321. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Gianaros PJ, Jennings JR. (2018) Host in the machine: A neurobiological perspective on psychological stress and cardiovascular disease . American Psychologist 73 ( 8 ): 1031–1044. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Golia E, Limongelli G, Natale F, et al. (2014) Inflammation and cardiovascular disease: From pathogenesis to therapeutic target . Current Atherosclerosis Reports 16 ( 9 ): 435. [ PubMed ] [ Google Scholar ]
- Gordon JL, Girdler SS, Meltzer-Brody SE, et al. (2015) Ovarian hormone fluctuation, neurosteroids, and HPA axis dysregulation in perimenopausal depression: A novel heuristic model . American Journal of Psychiatry 172 ( 3 ): 227–236. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Gouin J-P, Hantsoo L, Kiecolt-Glaser J. (2008) Immune dysregulation and chronic stress among older adults: A review . Neuroimmunomodulation 15 ( 4–6 ): 251–259. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Grivennikov SI, Greten FR, Karin M. (2010) Immunity, inflammation, and cancer . Cell 140 ( 6 ): 883–899. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Gross J. (2002) Emotion regulation: Affective, cognitive, and social consequences . Psychophysiology 39 : 281–291. [ PubMed ] [ Google Scholar ]
- Hamer M, Steptoe A. (2012) Cortisol responses to mental stress and incident hypertension in healthy men and women . The Journal of Clinical Endocrinology and Metabolism 97 ( 1 ): E29–E34. [ PubMed ] [ Google Scholar ]
- Hardt J, Rutter M. (2004) Validity of adult retrospective reports of adverse childhood experiences: Review of the evidence . Journal of Child Psychology and Psychiatry, and Allied Disciplines 45 ( 2 ): 260–273. [ PubMed ] [ Google Scholar ]
- Holt-Lunstad J, Smith TB, Baker M, et al. (2015) Loneliness and social isolation as risk factors for mortality . Perspectives on Psychological Science 10 ( 2 ): 227–237. [ PubMed ] [ Google Scholar ]
- Hughes ME, Waite LJ, Hawkley LC, et al. (2004) A short scale for measuring loneliness in large surveys: Results from two population-based studies . Research on Aging 26 ( 6 ): 655–672. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Kagan J. (2006) An overly permissive extension . Perspetives on Psychological Science 11 ( 4 ): 442–450. [ PubMed ] [ Google Scholar ]
- Karasek R. (1979) Job demands, job decision latitude, and mental strain: Implications for job redesign . Administrative Science Quarterly 24 ( 2 ): 285–308. [ Google Scholar ]
- Keyes KM, McLaughlin KA, Demmer RT, et al. (2013) Potentially traumatic events and the risk of six physical health conditions in a population-based sample . Depression and Anxiety 30 ( 5 ): 451–460. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Keys A, Longstreet Taylor H, Blackburn H, et al. (1971) Mortality and coronary heart disease among men studied for 23 years . Archives of Internal Medicine 128 ( 2 ): 201–214. [ PubMed ] [ Google Scholar ]
- Kiecolt-Glaser J, Glaser R, Shuttleworth EC, et al. (1987) Chronic stress and immunity in family caregivers of Alzheimer’s disease victims . Psychosomatic Medicine 49 ( 5 ): 523–535. [ PubMed ] [ Google Scholar ]
- Kirschbaum C, Pirke KM, Hellhammer DH. (1993) The ‘Trier social stress test’: A tool for investigating psychobiological stress responses in a laboratory setting . Neuropsychobiology 28 : 76–81. [ PubMed ] [ Google Scholar ]
- Knudsen EI. (2004) Sensitive periods in the development of the brain and behavior . Journal of Cognitive Neuroscience 16 ( 8 ): 1412–1425. [ PubMed ] [ Google Scholar ]
- Krause N, Shaw BA, Cairney J. (2004) A descriptive epidemiology of lifetime trauma and the physical health status of older adults . Psychology and Aging 19 ( 4 ): 637–648. [ PubMed ] [ Google Scholar ]
- Kudielka BM, Hellhammer DH, Wüst S. (2009) Why do we respond so differently? Reviewing determinants of human salivary cortisol responses to challenge . Psychoneuroendo-crinology 34 ( 1 ): 2–18. [ PubMed ] [ Google Scholar ]
- Landsbergis PA, Dobson M, Koutsouras G, et al. (2013) Job strain and ambulatory blood pressure: A meta-analysis and systematic review . American Journal of Public Health 103 ( 3 ): e61–e71. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- McArdle JJ, Epstein D. (1987) Latent growth curves within developmental structural equation models . Child Development 58 ( 1 ): 110–133. [ PubMed ] [ Google Scholar ]
- McEwen BS. (1998) Stress, adaptation, and disease: Allostasis and allostatic load . Annals of the New York Academy of Sciences 840 : 33–44. [ PubMed ] [ Google Scholar ]
- McEwen BS. (2015) The brain on stress: How behavior and the social environment “get under the skin.” In: Kaplan R, Spittel M, David DH. (eds) Population Health: Behavioral and Social Science Insights . Rockville, MD: Agency for Healthcare Research and Quality and Office of Behavioral and Social Sciences Research, National Institutes of Health, pp. 233–250. [ Google Scholar ]
- Madsen IEH, Nyberg ST, Magnusson Hanson LL, et al. (2017) Job strain as a risk factor for clinical depression: Systematic review and meta-analysis with additional individual participant data . Psychological Medicine 47 ( 8 ): 1342–1356. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Marsland AL, Walsh C, Lockwood K, et al. (2017) The effects of acute psychological stress on circulating and stimulated inflammatory markers: A systematic review and meta-analysis . Brain, Behavior, and Immunity 64 : 208–219. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Miller GE, Chen E, Cole SW. (2009) Health psychology: Developing biologically plausible models linking the social world and physical health . Annual Review of Psychology 60 ( 1 ): 501–524. [ PubMed ] [ Google Scholar ]
- Miller GE, Chen E, Parker KJ. (2011) Psychological stress in childhood and susceptibility to the chronic diseases of aging: Moving toward a model of behavioral and biological mechanisms . Psychological Bulletin 137 ( 6 ): 959–997. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Miller GE, Cohen S, Ritchey AK. (2002) Chronic psychological stress and the regulation of pro-inflammatory cytokines: A glucocorticoid-resistance model . Health Psychology 21 ( 6 ): 531–541. [ PubMed ] [ Google Scholar ]
- Muscatell KA, Eisenberger NI. (2012) A social neuroscience perspective on stress and health . Social and Personality Psychology Compass 6 : 890–904. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Nyberg ST, Fransson EI, Heikkilä K, et al. (2013) Job strain and cardiovascular disease risk factors: Meta-analysis of individual-participant data from 47,000 men and women . PLoS ONE 8 ( 6 ): e67323. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Pantell M, Rehkopf D, Jutte D, et al. (2013) Social isolation: A predictor of mortality comparable to traditional clinical risk factors . American Journal of Public Health 103 ( 11 ): 2056–2062. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Rohleder N. (2014) Stimulation of systemic low-grade inflammation by psychosocial stress . Psychosomatic Medicine 76 ( 3 ): 181–189. [ PubMed ] [ Google Scholar ]
- Rosengren A, Wilhelmsen L, Orth-Gomér K. (2004) Coronary disease in relation to social support and social class in Swedish men: A 15 year follow-up in the study of men born in 1933 . European Heart Journal 25 ( 1 ): 56–63. [ PubMed ] [ Google Scholar ]
- Roth DL, Fredman L, Haley WE. (2015) Informal caregiving and its impact on health: A reappraisal from population-based studies . Gerontologist 55 ( 2 ): 309–319. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Saxbe D, Rossin-Slater M, Goldenberg D. (2018) The transition to parenthood as a critical window for adult health . American Psychologist 73 ( 9 ): 1190–1200. [ PubMed ] [ Google Scholar ]
- Schulz R, Beach SR. (1999) Caregiving as a risk factor for mortality: The caregiver health effects study . JAMA 282 ( 23 ): 2215–2219. [ PubMed ] [ Google Scholar ]
- Selye H. (1956) The Stress of Life . New York: McGraw-Hill. [ Google Scholar ]
- Shiffman S, Stone AA, Hufford MR. (2008) Ecological momentary assessment . Annual Review of Clinical Psychology 4 ( 1 ): 1–32. [ PubMed ] [ Google Scholar ]
- Sin NL, Graham-Engeland JE, Ong AD, et al. (2015) Affective reactivity to daily stressors is associated with elevated inflammation . Health Psychology 34 ( 12 ): 1154–1165. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Slavich GM, Shields GS. (2018) Assessing lifetime stress exposure using the Stress and Adversity Inventory for Adults (Adult STRAIN): An overview and initial validation . Psychosomatic Medicine 80 : 17–27. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Slopen N, Koenen KC, Kubzansky LD. (2012) Childhood adversity and immune and inflammatory biomarkers associated with cardiovascular risk in youth: A systematic review . Brain, Behavior, and Immunity 26 ( 2 ): 239–250. [ PubMed ] [ Google Scholar ]
- Slopen N, Lewis TT, Gruenewald TL, et al. (2010) Early life adversity and inflammation in African Americans and whites in the midlife in the United States survey . Psychosomatic Medicine 72 ( 7 ): 694–701. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Slopen N, Meyer C, Williams D. (2018) Cumulative stress and health . In: Ryff CD, Krueger R. (eds) The Oxford Handbook of Integrative Health Science . Oxford: Oxford University Press, pp. 75–93. [ Google Scholar ]
- Steptoe A, Kivimäki M. (2013) Stress and cardiovascular disease: An update on current knowledge . Annual Review of Public Health 34 ( 1 ): 337–354. [ PubMed ] [ Google Scholar ]
- Steptoe A, Marmot M. (2005) Impaired cardiovascular recovery following stress predicts 3-year increases in blood pressure . Journal of Hypertension 23 ( 3 ): 529–536. [ PubMed ] [ Google Scholar ]
- Steptoe A, Kivimäki M, Lowe G, et al. (2016) Blood pressure and fibrinogen responses to mental stress as predictors of incident hypertension over an 8-year period . Annals of Behavioral Medicine 50 ( 6 ): 898–906. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Thoits P. (2010) Stress and health: Major findings and policy implications . Journal of Health and Social Behavior 51 ( S ): 41–53. [ PubMed ] [ Google Scholar ]
- Treiber FA, Kamarck T, Schneiderman N, et al. (2003) Cardiovascular reactivity and development of preclinical and clinical disease states . Psychosomatic Medicine 65 : 46–62. [ PubMed ] [ Google Scholar ]
- Van Den Bergh BRH, Mulder EJH, Mennes M, et al. (2005) Antenatal maternal anxiety and stress and the neurobehavioural development of the fetus and child: Links and possible mechanisms. A review . Neuroscience and Biobehavioral Reviews 29 : 237–258. [ PubMed ] [ Google Scholar ]
- Vitaliano PP, Zhang J, Scanlan JM. (2003) Is Caregiving Hazardous to One’s Physical Health? A Meta-Analysis . Psychological Bulletin 129 : 946–972. [ PubMed ] [ Google Scholar ]
- Weinstock M. (2001) Alterations induced by gestational stress in brain morphology and behaviour of the offspring . Progress in Neurobiology 65 : 427–451. [ PubMed ] [ Google Scholar ]
- Wheaton B, Young M, Montazer S, et al. (2013) Social stress in the twenty-first century . In: Aneshensel CS. (ed.) Handbook of the Sociology of Mental Health (2nd edn). Cham: Springer, pp. 299–323. [ Google Scholar ]
- Zeanah CH, Gunnar MR, McCall RB, et al. (2011) Sensitive periods . Monographs of the Society for Research in Child Development 76 ( 4 ): 147–162. [ PMC free article ] [ PubMed ] [ Google Scholar ]
- Research article
- Open access
- Published: 29 May 2024
Electroacupuncture attenuates neuropathic pain via suppressing BIP-IRE-1α-mediated endoplasmic reticulum stress in the anterior cingulate cortex
- Lin-Wei Ma 1 , 4 na1 ,
- Yu-Fan Liu 1 na1 ,
- Hui Zhang 1 ,
- Chang-Jun Huang 1 , 4 ,
- Xin-Zhe Qu 5 ,
- Jia-Piao Lin 1 ,
- Yan Yang 2 , 3 &
- Yong-Xing Yao ORCID: orcid.org/0000-0002-5458-8298 1
Biological Research volume 57 , Article number: 34 ( 2024 ) Cite this article
120 Accesses
1 Altmetric
Metrics details
Studies have suggested that endoplasmic reticulum stress (ERS) is involved in neurological dysfunction and that electroacupuncture (EA) attenuates neuropathic pain (NP) via undefined pathways. However, the role of ERS in the anterior cingulate cortex (ACC) in NP and the effect of EA on ERS in the ACC have not yet been investigated. In this study, an NP model was established by chronic constriction injury (CCI) of the left sciatic nerve in rats, and mechanical and cold tests were used to evaluate behavioral hyperalgesia. The protein expression and distribution were evaluated using western blotting and immunofluorescence. The results showed that glucose-regulated protein 78 (BIP) and inositol-requiring enzyme 1α (IRE-1α) were co-localized in neurons in the ACC. After CCI, BIP, IRE-1α, and phosphorylation of IRE-1α were upregulated in the ACC. Intra-ACC administration of 4-PBA and Kira-6 attenuated pain hypersensitivity and downregulated phosphorylation of IRE-1α, while intraperitoneal injection of 4-PBA attenuated hyperalgesia and inhibited the activation of P38 and JNK in ACC. In contrast, ERS activation by intraperitoneal injection of tunicamycin induced behavioral hyperalgesia in naive rats. Furthermore, EA attenuated pain hypersensitivity and inhibited the CCI-induced overexpression of BIP and pIRE-1α. Taken together, these results demonstrate that EA attenuates NP by suppressing BIP- and IRE-1α-mediated ERS in the ACC. Our study presents novel evidence that ERS in the ACC is implicated in the development of NP and provides insights into the molecular mechanisms involved in the analgesic effect of EA.
Neuropathic pain (NP) is caused by injury or disease of the somatosensory nervous system and commonly results from trauma, tumors, chemotherapy, and diabetes. NP is characterized by hyperalgesia, allodynia, and aberrant spontaneous pain [ 1 , 2 ]. NP has become a serious public health issue because of its high prevalence, debilitating effects, and high social costs [ 3 , 4 ]. Because the pathogenesis of NP is not yet fully understood, comprehensive treatments, such as drugs (e.g., tricyclic antidepressants, calcium channel blockers, opioids) and non-drug therapies (e.g., nerve stimulation), are commonly ineffective and often accompanied by serious side effects [ 5 , 6 ]. As a component of the limbic system, the anterior cingulate cortex (ACC) is mainly involved in the emotional-affective component of pain; however, increasing evidence has recently highlighted its important role in the modulation of the sensory-discriminative component of pain, although further exploration is necessary [ 7 , 8 ].
Endoplasmic reticulum stress (ERS) is a cellular protective process that is mediated by the ERS-sensing protein, pancreatic endoplasmic reticulum kinase (PERK), activating transcription factor 6 (ATF-6), inositol-requiring enzyme 1α (IRE-1α), and their downstream signaling molecules [ 9 , 10 ]. Under pathological conditions, the accumulation of unfolded proteins in the endoplasmic reticulum promotes the dissociation of ERS-sensing proteins from glucose regulatory protein 78 (BIP), activating the unfolded protein response and restoring cell homeostasis [ 11 , 12 ]. When hyperactivated by ERS, phosphorylated IRE-1α transitions from homodimers to high-order oligomers, inducing the activation or upregulation of numerous proinflammatory molecules, and participating in the development of multitudinous neurological diseases such as Alzheimer’s disease, Parkinson’s disease, and amyotrophic lateral sclerosis [ 13 , 14 ]. The role of BIP-IRE-1α-mediated ERS in the ACC in NP remains unclear.
Acupuncture is a traditional therapeutic technique that has been used in oriental medicine for approximately 3000 years. Since the 1970s, electroacupuncture (EA) has gradually replaced manual acupuncture because of its advantages, such as better analgesic effects and objectively quantified and controlled stimulation [ 15 , 16 ]. Currently, EA is widely used in stroke, Alzheimer’s disease, urinary incontinence, and other mental and physical disorders [ 17 , 18 , 19 ]. Recently, the analgesic effects of EA have been widely validated in basic and clinical studies [ 16 , 20 , 21 , 22 ]. However, the mechanism underlying the antihyperalgesic effects of EA requires further elucidation.
In the present study, we aimed to investigate the following: (1) the expression of BIP and IRE-1α and the ERS mediated by BIP-IRE-1α in the ACC, (2) its role in NP, and (3) whether the mechanisms underlying the antihyperalgesic effects of EA were associated with ERS mediated by BIP-IRE-1α in the ACC.
Cellular distribution of BIP and IRE-1α in the ACC
We first investigated the cell types expressing BIP and IRE-1α in the ACC using immunofluorescence experiments. Double immunofluorescence staining was performed for both BIP and IRE-1α using neuron-specific nuclear protein (NeuN), glial fibrillary acidic protein (GFAP), and ionized calcium-binding adaptor molecule-1 (Iba1). The results showed that BIP and IRE-1α co-localized well with NeuN but not with GFAP or Iba1 (Fig. 1 A, B).

Cellular distribution of BIP and IRE-1α in the anterior cingulate cortex. Double immunofluorescence staining showed colocalization of BIP (A) and IRE-1α (B) with NeuN, but not with GFAP or Iba1. Scale bar = 50 μm. BIP: Glucose-regulated protein 78, IRE-1α: Inositol-requiring enzyme 1α, NeuN: Neuron-specific nuclear protein, GFAP: Glial fibrillary acidic protein, Iba1: Ionized calcium-binding adaptor molecule-1
CCI induced behavioral hyperalgesia, upregulation of BIP and IRE-1α, and phosphorylation of IRE-1α in the ACC
A NP model was established by chronic constriction injury of the left sciatic nerve to explore the role of ERS in the ACC during NP. The MWT and ATS were determined before and 7 days after CCI, with no significant difference observed at baseline between the sham and CCI groups. Seven days after CCI, the MWT (Fig. 2 A) was significantly lower and the ATS (Fig. 2 B) was significantly higher in the CCI group than those in the sham group, demonstrating that CCI successfully induced behavioral hyperalgesia. Consistent with the behavioral hyperalgesia observed after CCI, the western blotting results showed that the expression of BIP (Fig. 2 C) and IRE-1α (Fig. 2 D) was bilaterally upregulated and the phosphorylation level of IRE-1α (Fig. 3 E) was bilaterally increased in the CCI group on day 7 after CCI. These data demonstrated that BIP-IRE-1α mediates ERS in the ACC after CCI.

CCI induced pain hypersensitivity, upregulation of BIP and IRE-1α, and phosphorylation of IRE-1α in the ACC. (A) Compared to the sham group, the CCI group showed a significantly lower MWT on the 7th day after surgery (*** p < 0.001, independent t- test, n = 7). (B) Compared to the sham group, the CCI group showed a significantly higher ATS on the 7th day after surgery (*** p < 0.001, independent t- test, n = 6). (C, D, E) Western blotting showed that compared to the sham group, the expression of BIP and IRE-1α in the ACC was bilaterally increased in the CCI group (*** p < 0.001, vs. sham, one-way ANOVA, n = 3); and the phosphorylation of IRE-1α was bilaterally increased in the CCI group (** p < 0.01, vs. sham, one-way ANOVA, n = 3). Error bars represent the standard error of the mean. CCI: Chronic constriction injury, BIP: Glucose-regulated protein 78, IRE-1α: Inositol-requiring enzyme 1α, ACC: Anterior cingulate cortex, MWT: Mechanical withdrawal threshold, ATS: Acetone test score
4-PBA and Kira-6 reversed hyperalgesia and inhibited CCI-induced IRE-1α activation
To further investigate the role of ERS in the ACC in the initiation of NP, the BIP inhibitor 4-PBA and the IRE-1α inhibitor Kira-6 were injected into the ACC from days 0 to 6 after CCI to suppress ERS in the ACC (Fig. 3 A, B, C). The MWT and ATS were measured before and 3, 5, and 7 days after CCI. The behavioral test results showed that the MWT and ATS did not differ significantly among the four groups before CCI; however, the MWT (Fig. 3 D) was significantly higher and the ATS (Fig. 3 E) was significantly lower in the CCI + 4-PBA and CCI + Kira-6 groups than those in the CCI + DMSO group. These results indicate that CCI-induced ERS plays an important role in the initiation of NP. Meanwhile, western blotting analysis showed that the expression level of BIP (Fig. 3 F) was not significantly different among the four groups, and the expression of IRE-1α (Fig. 3 G) and activation of IRE-1α (Fig. 3 H) were inhibited in the CCI + 4-PBA and CCI + Kira-6 groups, indicating successful suppression of ERS in the ACC. Furthermore, the results of western blot demonstrated a notable reduction in the expression levels of phosphorylated JNK (pJNK) (Fig. 3 I) and phosphorylated P38 (pP38) (Fig. 3 J) in the CCI + Kira-6 group, suggesting a link between the inhibition of ERS and the downregulation of P38 and JNK in the context of hyperalgesia.

The inhibitors 4-PBA and Kira-6 reversed hyperalgesia and inhibited CCI-induced activation of IRE-1α. (A, B, C) Schematic diagram and location of ACC microinjection. (D, E) Compared to the CCI + DMSO group, the MWT and ATS were not significantly different among the four groups before CCI; the MWT was significantly higher and the ATS was significantly lower in the CCI + 4-PBA group 5 and 7 days, and 3, 5, and 7 days after CCI in the CCI + Kira-6 group (*** p < 0.001, CCI + 4-PBA vs. CCI + DMSO; # p < 0.05, ## p < 0.01, ### p < 0.001, CCI + Kira-6 vs. CCI + DMSO, two-way ANOVA, n = 7). (F, G, H) The results of western blotting showed that the expression of BIP was not significantly different among the four groups; however, the expression and activation of IRE-1α and pIRE-1α were significantly inhibited in the CCI + 4-PBA and CCI + Kira-6 groups compared to those in the CCI + DMSO group (*** p < 0.001, one-way ANOVA, n = 3). (I, J) The western blotting analysis revealed a significant difference in the expression levels of pJNK and pP38 between the CCI + DMSO and CCI + Kira-6 groups (* p < 0.05, *** p < 0.001, CCI + Kira-6 vs. CCI + DMSO, one-way ANOVA, n = 3). Error bars represent the standard error of the mean. ACC: Anterior cingulate cortex, DMSO: Dimethyl sulfoxide, MWT: Mechanical withdrawal threshold, ATS: Acetone test score, BIP: Glucose-regulated protein 78, IRE-1α: Inositol-requiring enzyme 1α
Intraperitoneal injection of 4-PBA attenuated CCI-induced hyperalgesia
To further explore the influence of ERS on CCI-induced hyperalgesia from a translational perspective, intraperitoneal injections of 4-PBA were administered on the day of CCI and continued once a day for the following 7 days. The MWT and ATS were measured before CCI and on days 3, 5, and 7 after CCI. The results revealed a significant increase in the MWT (Fig. 4 A) and a substantial reduction in the ATS (Fig. 4 B) in the CCI + 4-PBA group, with no significant impact on locomotive ability (Fig. 4 C), as compared to the CCI + DMSO group. Furthermore, western blotting analysis indicated that BIP expression was inhibited in the CCI + 4-PBA group (Fig. 4 D). Although the expression level of IRE-1α was not significantly different between the three groups (Fig. 4 E), the activation of pIRE-1α was inhibited in the CCI + 4-PBA group (Fig. 4 F). These data suggest that the inhibition of ERS might alleviate CCI-induced hyperalgesia without affecting locomotive ability, the underlying mechanisms of which warrant further investigation.

Intraperitoneal injection of 4-PBA reversed hyperalgesia induced by CCI and suppressed CCI-triggered BIP expression and IRE-1α activation. (A, B) Compared to the CCI + DMSO group, the MWT and ATS were not significantly different in the CCI + 4-PBA before CCI; the MWT was significantly higher on days 5 and 7, and the ATS was significantly lower on days 3, 5, and 7 in the CCI + 4-PBA group after CCI (** p < 0.01, *** p < 0.001, CCI + 4-PBA vs. CCI + DMSO; n = 6). (C) The strength of grasp of the front paws showed no significant differences among the CCI, CCI + DMSO, and CCI + 4-PBA groups ( n = 6). (D, E, F) The expression of IRE-1α was not significantly different among the three groups, but that of BIP and pIRE-1α was significantly decreased in the CCI + 4-PBA group (* p < 0.05, ** p < 0.01, one-way ANOVA, n = 3). Error bars represent the standard error of the mean. CCI: Chronic constriction injury, BIP: Glucose-regulated protein 78, IRE-1α: Inositol-requiring enzyme 1α, MWT: Mechanical withdrawal threshold, ATS: Acetone test score
Tunicamycin-induced hyperalgesia in naive rats
To investigate whether ERS induction could lead to hypersensitivity, we next administered intraperitoneal injections of tunicamycin, a known activator of ERS, into naive rats. The MWT and ATS were assessed before and 2 and 24 h after injection. The behavioral results demonstrated that, compared to the DMSO group, the tunicamycin group exhibited a substantial decrease in MWT (at both 2 and 24 h), and a significant increase in ATS (2 h) after application (Fig. 5 A, B). These findings indicate that ERS activation induces hyperalgesia in naive rats.

Tunicamycin injection induces hyperalgesia in naive rats. The tunicamycin group exhibited significantly lower mechanical withdrawal threshold (A, 2 and 24 h) and higher acetone test score (B, 2 h) than the DMSO group post-injection (* p < 0.05, *** p < 0.001, two-way ANOVA, n = 6). Error bars represent the standard error of the mean. DMSO: Dimethyl sulfoxide
EA attenuated hyperalgesia and suppressed CCI-induced ERS in ACC
To further investigate the antihyperalgesic effect and mechanisms of EA, the consecutive EA procedure was applied from days 0 to 6 after CCI for 30 min/day. The MWT and ATS were measured before and on days 3, 5, and 7 after CCI. The behavioral results showed that the MWT and ATS were not significantly different among the four groups before and 3 days after CCI; however, the MWT (Fig. 6 A) was significantly higher and the ATS (Fig. 6 B) was significantly lower in the CCI + EA group than in the CCI + SEA group, indicating that EA inhibits the initiation of NP. Further, western blotting showed that the expression of BIP was significantly lower in the CCI + EA group than that in the CCI + SEA group (Fig. 6 C). The expression of IRE-1α was not significantly different among the three groups (Fig. 6 D), but the phosphorylation of IRE-1α was significantly decreased in the CCI + EA group compared to that in the CCI + SEA group (Fig. 6 E). These findings demonstrate that the EA attenuates NP by suppressing ERS in the ACC after CCI.

Electroacupuncture attenuated hyperalgesia and suppressed CCI-induced endoplasmic reticulum stress in the anterior cingulate cortex. (A, B) Compared to the CCI + SEA group, the MWT and ATS showed no significant differences among the three groups before and 3 days after CCI. However, the MWT was significantly higher and the ATS was significantly lower in the CCI + EA group on days 5 and 7 after CCI (** p < 0.01, *** p < 0.001, CCI + EA vs. CCI + SEA, two-way ANOVA, n = 7). (C) Compared to the CCI + SEA group, the expression of BIP was significantly lower in the CCI + EA group (* p < 0.05, one-way ANOVA, n = 5). (D, E) The expression of IRE-1α was not significantly different among the three groups, but the phosphorylation of IRE-1α was significantly decreased in the CCI + EA group (** p < 0.01, one-way ANOVA, n = 5). Error bars represent the standard error of the mean. CCI: Chronic constriction injury, SEA: sham EA, EA: electroacupuncture, BIP: Glucose-regulated protein 78, IRE-1α: Inositol-requiring enzyme 1α, MWT: Mechanical withdrawal threshold, ATS: Acetone test score
Here, we demonstrated that both BIP and IRE-1α are expressed in neurons but not in astrocytes or microglia in the ACC. Additionally, we showed that the abundance of BIP and IRE-1α bilaterally increased, while IRE-1α was activated by phosphorylation 7 days after CCI surgery, consistent with the corresponding CCI-induced behavioral hypersensitivity. Intra-ACC microinjection or intraperitoneal injection of the BIP or IRE-1α inhibitor impeded the initiation of CCI-induced NP. Activation of ERS induced hypersensitivity in naive rats. These results indicate that BIP-IRE-1α-mediated ERS in the ACC contributes to peripheral nerve injury-induced NP. Furthermore, EA attenuated hyperalgesia and suppressed BIP overexpression and the activation of IRE-1α during CCI-induced ERS in the ACC, suggesting that EA hampered NP initiation by suppressing ERS in the ACC.
The ACC, a highly heterogeneous cortex with extensive fiber connections, receives and integrates afferent information from other regions of the central nervous system. The ACC has been recognized to play a critical role in the emotional-affective component [ 23 , 24 ] and has recently emerged as a region involved in modulating the sensory-discriminative component of pain [ 23 ]. Indeed, photogenetic studies have demonstrated that specific activation of vertebral neurons in the ACC reduces the threshold of mechanical stimuli in mice [ 25 ]. Meanwhile, the activation of inhibitory interneurons significantly inhibits nociceptive perception [ 26 ], whereas pharmacological inhibition of chemokine receptor 3 (CXCR3) in the ACC significantly relieves hyperalgesia in NP rats [ 27 ]. This evidence suggests that there may be a specific neural circuit involved in nociceptive perception in the ACC, although whether ERS in the ACC is involved in NP needs to be further explored.
Previous studies have shown that ERS plays an important role in the development of neurodegenerative diseases, cerebral ischemia, spinal cord injury, multiple sclerosis, and diabetic neuropathy [ 28 , 29 , 30 , 31 ]. Furthermore, the BIP-IRE-1α pathway is activated in the peripheral nervous system and spinal cord under NP [ 10 , 32 , 33 ], and local or systemic use of ERS inhibitors could significantly relieve hyperalgesia [ 34 , 35 ], indicating the important role of BIP-IRE-1α pathway-mediated ERS in NP. However, whether ERS could be triggered in the ACC, and its role in NP have not yet been explored. In the current study, the NP model was established using CCI to investigate the role of ERS in ACC in the sensory-discriminative component of pain. Immunofluorescence analysis revealed that both BIP and IRE-1α were co-localized with NeuN, which was consistent with the findings of a previous study [ 35 , 36 ], providing the theoretical basis for ERS in the ACC. Next, western blotting was used to detect protein expression to further investigate the role of BIP-IRE-1α-mediated ERS in pain modulation. The results showed bilateral upregulation of BIP and IRE-1α in the ACC after CCI, while IRE-1α was bilaterally activated in a phosphorylated manner, unlike the unilateral activation observed in the spinal cord [ 35 ]. This discrepancy may be attributed to the crossover of the spinothalamic tracts in the spinal cord and afferent projections from the contralateral pain-transmitting area [ 37 , 38 ]. This result indicated the induction of BIP-IRE-1α pathway-mediated ERS after CCI in the ACC. To further explore the role of BIP-IRE-1α-mediated ERS in the ACC during pain modulation, the BIP inhibitor, 4-PBA, and the IRE-1α inhibitor, Kira-6, were consecutively administered via ACC microinjection and intraperitoneal injection respectively to inhibit CCI-induced ERS. Behavioral tests revealed that both 4-PBA and Kira-6 impeded the initiation of mechanical hyperalgesia and cold allodynia after CCI, suggesting a critical role for ERS in the ACC in CCI-induced NP. Interestingly, there were no significant behavioral differences among the four groups on day 3 after CCI, but significant differences were observed on days 5 and 7. This phenomenon may be attributed to the delayed action of the drugs, or incision-mediated nociceptive hyperactivity that was not modulated by ERS in the ACC. However, the exact mechanism underlying this phenomenon requires further investigation. In line with the results of previous studies [ 34 ], the results of western blotting showed that both 4-PBA and Kira-6 significantly suppressed the expression of IRE-1α and its phosphorylation, indicating successful inhibition of ERS in the ACC.
JNK and p38 are well-recognized kinases involved in pain modulation. We analyzed pJNK and pP38 expression using western blotting, with the results showing a significant decrease in expression within the ACC of rats administered Kira-6. This reduction implies a potential connection between the inhibition of ERS and the downregulation of P38 and JNK, which may contribute to reduced production of inflammatory cytokines.
The suppression of ERS has potential side effects, including disruptions in the intricate process of protein folding, disturbances in the regulation of intracellular calcium, and modifications to the unfolded protein response pathway [ 39 ]. In the present study, we evaluated locomotor ability using forelimb grip strength, but found no significant differences in limb strength between the rats treated with inhibitors and the control group, suggesting that the application of ER inhibitors does not impair the locomotive ability of rats. However, given the limitations inherent to animal studies, other side effects, such as abnormal sensations, could not be ruled out.
Previous studies have confirmed that the mechanisms underlying the analgesic effect of EA are via the regulation of opioids, norepinephrine, or cholinergic activity in subcortical structures such as the spinal cord, periaqueductal gray matter, and hypothalamus pre-optic area [ 16 , 40 , 41 , 42 ]. However, whether the cerebral cortex is involved in the antihyperalgesic effects of EA remains unclear. By regulating ERS, EA can alleviate neuronal apoptosis caused by ischemic stroke or spinal cord injury and promote the recovery of nerve function [ 43 , 44 ], providing a theoretical possibility for EA to alleviate NP by suppressing ERS. In the present study, we performed EA treatment from days 0 to 6 for 30 min/day. The behavioral results showed no significant differences between the baseline and day 3 after CCI; however, the MWT was significantly increased and the ATS was significantly decreased in the CCI-EA group on days 5 and 7 after CCI, indicating that consecutive EA treatment effectively hampered the initiation of NP after CCI. Similarly, the delayed antihyperalgesic effect of EA might be attributed to the delayed action of the EA, or incision-induced nociceptive hyperactivity and acute nervous inflammation were not modulated by ERS in the ACC. Western blotting was used to further elucidate the mechanisms underlying the antihyperglycemic effect of EA, with the results showing that EA inhibited the overexpression of BIP and the CCI-induced activation of IRE-1α, suggesting that EA attenuated NP by suppressing BIP-IRE-1α-mediated ERS in the ACC (Fig. 7 ).

An explanatory graphic illustrating electroacupuncture attenuates neuropathic pain by suppressing endoplasmic reticulum stress in the anterior cingulate cortex. ERS: Endoplasmic reticulum stress, ACC: Anterior cingulate cortex, UPR: Unfolded protein response
Limitations
Our study has limitations that warrant discussion. First, although we have revealed that the BIP-IRE-1α pathway in the ACC was activated after peripheral nerve injury, that both the antagonists of BIP and IRE-1α mitigated CCI-induced NP, and that the activation of this pathway induces pain hypersensitivity, the mechanisms underlying ERS mediating NP were not well-addressed. Second, only one dose of inhibitors was tested in the present study, and we do not know whether the effect is dose-dependent or not. Third, studies demonstrated that IRE-1α-, ATF6-, and PERK-mediated ERS played an important role in the development of neuropathic pain in the spinal cord [ 35 ], but the role of ATF6 and PERK in the ACC during NP and whether they are involved in the high central mechanisms of the antihyperalgesic effects of EA need to be further elucidated.
Conclusions
The present study demonstrated that BIP-IRE-1α-mediated ERS in the ACC plays a critical role in the initiation of NP and that EA suppresses the initiation of NP induced by peripheral nerve injury by suppressing BIP-IRE-1α-mediated ERS in the ACC. We present novel evidence that ERS in the ACC is implicated in the development of NP and provide insight into the molecular mechanisms involved in the analgesic effect of EA.
Materials and methods
Male Sprague–Dawley rats (6–8 weeks old; weight, 220 ± 20 g) were obtained from the Experimental Animal Center of the Zhejiang Academy of Medical Sciences. Rats were housed in groups (3–4 per cage) at a temperature of 24 ± 2 °C under a 12-h light/dark cycle, with food and water provided ad libitum . The rats were randomly allocated to each group and allowed a week to adapt to the new environment before initiating the experiment. All animal experiments complied with the ARRIVE guidelines, internationally accredited guidelines, and ethical regulations on animal research [ 45 ]. The study protocol was approved by the Research Ethics Committee of the First Affiliated Hospital of Zhejiang University. All efforts were made to minimize the number of animals used and their suffering. The experimental timeline used in this study is shown in Fig. 8 .

Illustration of the experimental timelines
Induction of NP
After baseline had been obtained, the rats were randomly assigned to the sham and chronic constriction injury (CCI) groups. Under isoflurane anesthesia, as reported by Bennett and Xie and in our previous studies [ 46 , 47 , 48 ], the left sciatic nerve of the rat was exposed through blunt dissection of the middle thigh. In the CCI group, the sciatic nerve was isolated and ligated using three strands of 4 − 0 chromic gut sutures (Pudong Jinhuan Co. Ltd., Shanghai, China) placed 1 mm apart. The muscles and skin were closed layer-by-layer with 4 − 0 sutures. In the sham group, the left sciatic nerve was visualized but not ligated. After surgery, the rats were subcutaneously injected with 80,000 U of penicillin to prevent infection.
Behavioral tests
The rats were allowed to acclimate for 3 consecutive days (30 min per day) in a plastic box (12 cm × 15 cm × 22 cm) on an elevated wire mesh before behavioral tests. The experimenter was blinded to the treatment received by the rats.
Mechanical withdrawal threshold
The mechanical withdrawal threshold (MWT) was determined using a set of von Frey filaments. Briefly, the left plantar surface was stimulated with filaments of increasing stiffness (0.4–26 g) until a quick withdrawal or licking of the paw was noted, and the magnitude of the filaments was recorded as the MWT. The testing was repeated three times with an interval of 5 min, and the average value was considered the final MWT.
Acetone test score
Cold allodynia was tested using the acetone test score (ATS), as described by Farsi et al. [ 49 ]. on the same apparatus as the MWT test. Briefly, 100 µL of acetone was sprayed onto the left plantar surface, and the responses were observed for 20 s after application. The results were scored on a 4-point scale as follows: 0, no response; 1, startle response without paw withdrawal; 2, brief withdrawal of the paw; 3, prolonged withdrawal (5–30 s); and 4, prolonged and repetitive withdrawal along with flinching and/or licking. The testing was repeated three times at 5-min intervals, and the average value was considered as the final ATS.
ACC catheterization and drug administration
For intra-ACC drug administration, ACC catheterization was performed under anesthesia with pentobarbital sodium (60 mg/kg) according to our previous report [ 27 ]. Briefly, the head was fixed on a stereotactic apparatus in the prone position, and a 1-cm longitudinal incision was made in the middle of the head to expose the skull. Two holes were drilled on each side (Bregma forward, 1.7 mm; lateral, 0.6 mm) and a trocar (Shenzhen Ruiwode Life Science and Technology Co., Ltd, Guangdong, China) was inserted. Two small screws were installed superficially in the occipital bone, dental methyl methacrylate was used to fix the trocar with screws, and the rats were administered a subcutaneous injection of 80,000 U penicillin to prevent infection. CCI was performed 7 days after catheterization. The BIP and IRE-1 inhibitors 4-PBA and Kira-6 were purchased from Selleck (Houston, TX, USA) and dissolved in 10% dimethyl sulfoxide (DMSO). The rats were randomly assigned to the following four groups: CCI, CCI + DMSO, CCI + 4-PBA, and CCI + Kira-6. Both 4-PBA and Kira-6 were dissolved in 10% DMSO at a concentration of 20 µg/µL. Drugs or vehicle were bilaterally injected into the ACC (1 µL per side) from days 0 to 6 after CCI surgery.
Systemic administration
For systemic administration, intraperitoneal injections of 4-PBA (dissolved in 10% DMSO) were administered on the day of CCI and continued once a day for the following 7 days at a dosage of 20 mg/kg.
For ERS activation, a single intraperitoneal injection of tunicamycin, a recognized activator of ERS, was administered to naive rats. Tunicamycin was obtained from Aladdin Reagent (Shanghai, China) and dissolved in 10% DMSO. The rats were randomly assigned to the naive, DMSO (10% DMSO, 1 mL), and tunicamycin (2 µg/kg, in a 1 mL volume) groups. All animals underwent behavioral testing before and 2 and 24 h after administration.
Locomotive ability
We used the YLS-13 A grasp tester (Jinan Yiyan Science Co. Ltd., Shandong, China) to measure the grip force of the anterior claw and evaluate locomotive ability. The experimental protocol involved horizontally situating the grip tester on the ground, placing the rat onto a flat plate, and firmly fastening the front paw onto the steel wire. The incremental force was gradually applied against gravity until the front paw released its grip on the wire. Subsequently, the grip force was automatically recorded. During each session, three measurements were collected at 5-min intervals, and the average was computed to determine the final grip force.
- Electroacupuncture
The EA procedure was performed according to the methods outlined in a previous study [ 50 ]. After baseline was obtained, the rats were randomly allocated to the following three groups: CCI, CCI + sham EA (CCI + SEA), and CCI + EA (CCI + EA). Briefly, acupuncture needles (diameter, 0.25 mm; depth, 4 mm) were inserted into the left Zusanli (ST36, 5 mm lateral to the anterior tubercle of the tibia) and left Kunlun (BL60; the sunken area between the lateral malleolus and the Achilles tendon) acupoints. The needles were connected to a HuaTuo acupuncture nerve stimulator (HuaTuo-SDZ-II; Suzhou Medical Appliance Co., Ltd., Suzhou, Jiangsu). The EA parameters were set as follows: 2 Hz, consecutive wave output, which lasted for 30 min, with intensities ranging from 0.5 to 1.5 mA (increased by 0.5 mA every 10 min). For sham EA treatment, needles were inserted as in the EA group, but without electrical stimulation. EA treatment was performed once daily for 7 consecutive days, from days 0 to 6 after CCI.
Immunofluorescence assay
After deep anesthesia with pentobarbital sodium, the rats were transcardially perfused with 150 mL of 1 × phosphate buffered saline (PBS) (4 °C), followed by 150 mL of 4% paraformaldehyde (4 °C). The ACC was harvested, fixed with 4% paraformaldehyde for 48 h, and then dehydrated with 30% sucrose for 3 days at 4 °C. Subsequently, the ACC was transversely cut into slices (30-µm thick). The sections were blocked with 10% sheep or donkey serum for 2 h at room temperature and incubated with the following primary antibodies for 48 h at 4 °C: goat-anti-Iba1 (1:200, Abcam, Cambridge, UK), mouse anti-GFAP (1:500, Cell Signaling Technology), mouse anti-NeuN (1:2000, Abcam), rabbit anti-BIP (1:500, Cell Signaling Technology), and rabbit-anti-IRE-1 (1:1000, Proteintech). The sections were washed with 1 × PBS and incubated with fluorescent secondary antibodies in the dark for 2 h at room temperature. Finally, the sections were examined under a fluorescence microscope (FV3000; Olympus, Tokyo, Japan).
Western blot analysis
After deep anesthesia with pentobarbital sodium, the rats were decapitated, the brain was harvested, and the ACC was divided into the left (Ipsi.) and right (Contra.) sections as described in our previous study [ 27 ]. The ipsilateral protein samples were separated by SDS-PAGE and transferred to a PVDF membrane. Subsequently, the membranes were blocked in 5% skim milk at room temperature for 1 h before incubation with the following primary antibodies for 24 h at 4 °C: rabbit-anti-BIP (1:500, Cell Signaling Technology, Danvers, MA, USA), rabbit-anti-IRE-1 (1:1000, Proteintech, Rosemont, IL, USA), rabbit-anti-pIRE-1 (1:1000, Proteintech), and mouse-anti-GAPDH (1:10,000, Proteintech). After washing with TBST, the membrane was incubated with an HRP-conjugated secondary antibody for 2 h at room temperature. The ChemiDoc MP System (Bio-Rad, Hercules, CA, USA) was used to detect complex immune bands.
Statistical analysis
All data are expressed as the mean ± SEM and were analyzed using GraphPad Prism 8.0 (GraphPad, San Diego, CA, USA). Behavioral data were compared between the two groups using independent t -tests. The results of behavioral tests across different time points were analyzed using repeated measures two-way analysis of variance (ANOVA), followed by Bonferroni’s post hoc test. Western blot data were analyzed using one-way ANOVA, followed by Tukey’s multiple comparisons test. Statistical significance was set at p < 0.05.
Data availability
The datasets generated during and/or analyzed during the study are available from the corresponding author on reasonable request.
Abbreviations
- Neuropathic pain
- Endoplasmic reticulum stress
- Anterior cingulate cortex
Chronic constriction injury
Glucose-regulated protein 78
Inositol-requiring enzyme 1α
Analysis of variance
Neuron-specific nuclear protein
Glial fibrillary acidic protein
Ionized calcium-binding adaptor molecule-1
Fitzcharles MA, et al. Nociplastic pain: towards an understanding of prevalent pain conditions. Lancet. 2021;397(10289):2098–110.
Article PubMed Google Scholar
Kremer M, et al. Antidepressants and gabapentinoids in neuropathic pain: mechanistic insights. Neuroscience. 2016;338:183–206.
Article CAS PubMed Google Scholar
Cohen SP, Mao J. Neuropathic pain: mechanisms and their clinical implications. BMJ. 2014;348:f7656.
Finnerup NB, et al. Neuropathic pain: an updated grading system for research and clinical practice. Pain. 2016;157(8):1599–606.
Article CAS PubMed PubMed Central Google Scholar
Finnerup NB, Kuner R, Jensen TS. Neuropathic Pain: from mechanisms to treatment. Physiol Rev. 2021;101(1):259–301.
Wang Z, et al. Altered thalamic neurotransmitters metabolism and functional connectivity during the development of chronic constriction injury induced neuropathic pain. Biol Res. 2020;53(1):36.
Li XH, et al. Oxytocin in the anterior cingulate cortex attenuates neuropathic pain and emotional anxiety by inhibiting presynaptic long-term potentiation. Cell Rep. 2021;36(3):109411.
Xiao X, Zhang YQ. A new perspective on the anterior cingulate cortex and affective pain. Neurosci Biobehav Rev. 2018;90:200–11.
Liu Q, et al. Endoplasmic reticulum stress in autoimmune diseases. Immunobiology. 2020;225(2):151881.
Zhang HY, et al. Endoplasmic reticulum stress: relevance and therapeutics in central nervous system diseases. Mol Neurobiol. 2015;51(3):1343–52.
Kaneko M, et al. ER stress and disease: toward Prevention and Treatment. Biol Pharm Bull. 2017;40(9):1337–43.
Mao Y, et al. Endoplasmic reticulum stress contributes to Nociception via Neuroinflammation in a murine bone Cancer Pain Model. Anesthesiology. 2020;132(2):357–72.
Ghemrawi R, Khair M. Endoplasmic reticulum stress and unfolded protein response in neurodegenerative diseases. Int J Mol Sci, 2020. 21(17).
Oakes SA, Papa FR. The role of endoplasmic reticulum stress in human pathology. Annu Rev Pathol. 2015;10:173–94.
Qiu ZY et al. The Expression Patterns of c-Fos and c-Jun Induced by Different Frequencies of Electroacupuncture in the Brain Evid Based Complement Alternat Med, 2015. 2015: p. 343682.
Zhang R, et al. Mechanisms of acupuncture-electroacupuncture on persistent pain. Anesthesiology. 2014;120(2):482–503.
Comachio J, et al. Efficacy of acupuncture and electroacupuncture in patients with nonspecific low back pain: study protocol for a randomized controlled trial. Trials. 2015;16:469.
Article PubMed PubMed Central Google Scholar
Liu Z, et al. Effect of electroacupuncture on urinary leakage among women with stress urinary incontinence: a Randomized Clinical Trial. JAMA. 2017;317(24):2493–501.
Song YY, et al. Mechanisms of Electroacupuncture on Alzheimer’s disease: a review of Animal studies. Chin J Integr Med. 2020;26(6):473–80.
Jiang M, et al. Electroacupuncture suppresses glucose metabolism and GLUT-3 expression in medial prefrontal cortical in rats with neuropathic pain. Biol Res. 2021;54(1):24.
Kong JT, et al. Effect of Electroacupuncture vs Sham Treatment on Change in Pain Severity among adults with chronic low back Pain: a Randomized Clinical Trial. JAMA Netw Open. 2020;3(10):e2022787.
Lv ZT, et al. Effects of intensity of electroacupuncture on chronic pain in patients with knee osteoarthritis: a randomized controlled trial. Arthritis Res Ther. 2019;21(1):120.
Bliss TV, et al. Synaptic plasticity in the anterior cingulate cortex in acute and chronic pain. Nat Rev Neurosci. 2016;17(8):485–96.
Li Y. Synaptic plasticity and synchrony in the Anterior Cingulate Cortex Circuitry: a neural Network Approach to Causality of Chronic Visceral Pain and Associated Cognitive deficits. Adv Neurobiol. 2018;21:219–45.
Kang SJ, et al. Bidirectional modulation of hyperalgesia via the specific control of excitatory and inhibitory neuronal activity in the ACC. Mol Brain. 2015;8(1):81.
Gu L, et al. Pain inhibition by optogenetic activation of specific anterior cingulate cortical neurons. PLoS ONE. 2015;10(2):e0117746.
Qin J, et al. CXCR3 contributes to neuropathic pain via ERK activation in the anterior cingulate cortex. Biochem Biophys Res Commun. 2020;531(2):166–71.
Hashimoto S, et al. Endoplasmic reticulum stress responses in mouse models of Alzheimer’s disease: overexpression paradigm versus knockin paradigm. J Biol Chem. 2018;293(9):3118–25.
Kroeger H, et al. ER stress and unfolded protein response in ocular health and disease. FEBS J. 2019;286(2):399–412.
Medinas DB, et al. Endoplasmic reticulum stress leads to accumulation of wild-type SOD1 aggregates associated with sporadic amyotrophic lateral sclerosis. Proc Natl Acad Sci U S A. 2018;115(32):8209–14.
Peng T, et al. Fluoxetine-mediated inhibition of endoplasmic reticulum stress is involved in the neuroprotective effects of Parkinson’s disease. Aging. 2018;10(12):4188–96.
Inceoglu B, et al. Endoplasmic reticulum stress in the peripheral nervous system is a significant driver of neuropathic pain. Proc Natl Acad Sci U S A. 2015;112(29):9082–7.
Yang ES, et al. Involvement of endoplasmic reticulum stress response in orofacial inflammatory pain. Exp Neurobiol. 2014;23(4):372–80.
Chopra S et al. IRE1alpha-XBP1 signaling in leukocytes controls prostaglandin biosynthesis and pain. Science, 2019. 365(6450).
Liu Y, et al. Dexmedetomidine alleviated endoplasmic reticulum stress via inducing ER-phagy in the spinal cord of Neuropathic Pain Model. Front Neurosci. 2020;14:90.
Zhang E, et al. Endoplasmic reticulum stress impairment in the spinal dorsal horn of a neuropathic pain model. Sci Rep. 2015;5:11555.
Giesler GJ Jr., et al. Spinothalamic tract neurons that project to medial and/or lateral thalamic nuclei: evidence for a physiologically novel population of spinal cord neurons. J Neurophysiol. 1981;46(6):1285–308.
Granum SL. The spinothalamic system of the rat. I. locations of cells of origin. J Comp Neurol. 1986;247(2):159–80.
Grandjean JMD, et al. Pharmacologic IRE1/XBP1s activation confers targeted ER proteostasis reprogramming. Nat Chem Biol. 2020;16(10):1052–61.
Chen Y, et al. Neuronal GRK2 regulates microglial activation and contributes to electroacupuncture analgesia on inflammatory pain in mice. Biol Res. 2022;55(1):5.
Huang CP, et al. Electroacupuncture relieves CCI-Induced Neuropathic Pain Involving Excitatory and Inhibitory neurotransmitters. Evid Based Complement Alternat Med. 2019;2019:p6784735.
Article Google Scholar
Zhang XH, et al. Electroacupuncture attenuates Neuropathic Pain and Comorbid negative behavior: the involvement of the dopamine system in the Amygdala. Front Neurosci. 2021;15:657507.
Dai P, et al. [Effects of electroacupuncture at Jiaji(EX-B2) on autophagy and endoplasmic reticulum stress in spinal cord injury mice]. Zhen Ci Yan Jiu. 2021;46(1):45–51.
PubMed Google Scholar
Zhang YM, et al. Electroacupuncture regulates endoplasmic reticulum stress and ameliorates neuronal Injury in rats with Acute ischemic stroke. Evid Based Complement Alternat Med. 2021;2021:p9912325.
Google Scholar
Zimmermann M. Ethical guidelines for investigations of experimental pain in conscious animals. Pain. 1983;16(2):109–10.
Bennett GJ, Xie YK. A peripheral mononeuropathy in rat that produces disorders of pain sensation like those seen in man. Pain. 1988;33(1):87–107.
Li HL, et al. C-X-C motif chemokine 10 contributes to the Development of Neuropathic Pain by increasing the permeability of the blood-spinal cord barrier. Front Immunol. 2020;11:477.
Lin JP, et al. Dexmedetomidine Attenuates Neuropathic Pain by Inhibiting P2X7R Expression and ERK Phosphorylation in Rats. Exp Neurobiol. 2018;27(4):267–76.
Farsi L, et al. Postinjury treatment with magnesium sulfate attenuates neuropathic pains following spinal cord injury in male rats. Behav Pharmacol. 2015;26(3):315–20.
Li Y et al. Electroacupuncture alleviates Paclitaxel-Induced Peripheral Neuropathic Pain in rats via suppressing TLR4 signaling and TRPV1 upregulation in sensory neurons. Int J Mol Sci, 2019. 20(23).
Download references
Acknowledgements
Not applicable.
This research was supported by the Science and Technology Innovation Program of China-2030 (2021ZD0200408 [Yang]), National Natural Science Foundation of Zhejiang Province, China (grants LZ21H090001 and LBY22H270007 [Yao]), Zhejiang Provincial Outstanding Youth Science Foundation (LR23C090001 [Yang]), and the National Natural Science Foundation of China (grant 32071019 [Yang]).
Author information
Lin-Wei Ma and Yu-Fan Liu contributed equally to this work.
Authors and Affiliations
Department of Anesthesia, First Affiliated Hospital, Zhejiang University School of Medicine, 79 Qingchun Road, Hangzhou, 310003, China
Lin-Wei Ma, Yu-Fan Liu, Hui Zhang, Chang-Jun Huang, Ang Li, Jia-Piao Lin & Yong-Xing Yao
Department of Neurobiology, Sir Run Run Shaw Hospital, Zhejiang University School of Medicine, 3 East Qingchun Road, Hangzhou, 310020, China
School of Brain Science and Brain Medicine, Zhejiang University, Hangzhou, 310058, China
Department of Anesthesia, First People’s Hospital of Linping District of Hangzhou City, 369 Yingbin Road, Hangzhou, 311100, China
Lin-Wei Ma & Chang-Jun Huang
Department of Orthopedics, Graduated School, Xuzhou Medical University, 209 Tongshan Road, Xuzhou, 221004, China
You can also search for this author in PubMed Google Scholar
Contributions
L-WM: Conceptualization, Methodology, Behavioral test, performed western blotting, Formal analysis, Writing – original draft. Y-FL: Methodology, Behavioral tests, Formal analysis, Writing – review & editing. HZ: Methodology, Validation. C-JH: Performed western blotting. AL: Performed immunofluorescence. X-ZQ: Performed western blotting. J-PL: Provided technical support. YY: Writing – review & editing, Supervision, Funding acquisition. Y-XY: Conceptualization, Writing – review & editing, Supervision, Funding acquisition.
Corresponding authors
Correspondence to Yan Yang or Yong-Xing Yao .
Ethics declarations
Ethics approval and consent to participate.
All animal experiments complied with the ARRIVE guidelines and were approved by the Research Ethics Committee of the First Affiliated Hospital of Zhejiang University (No. 2019–232).
Consent for publication
Conflict of interest.
The authors declare that they have no competing financial interests or potential conflicts of interest.
Additional information
Publisher’s note.
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplementary Material 1
Rights and permissions.
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/ . The Creative Commons Public Domain Dedication waiver ( http://creativecommons.org/publicdomain/zero/1.0/ ) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
Reprints and permissions
About this article
Cite this article.
Ma, LW., Liu, YF., Zhang, H. et al. Electroacupuncture attenuates neuropathic pain via suppressing BIP-IRE-1α-mediated endoplasmic reticulum stress in the anterior cingulate cortex. Biol Res 57 , 34 (2024). https://doi.org/10.1186/s40659-024-00511-3
Download citation
Received : 14 November 2022
Accepted : 30 April 2024
Published : 29 May 2024
DOI : https://doi.org/10.1186/s40659-024-00511-3
Share this article
Anyone you share the following link with will be able to read this content:
Sorry, a shareable link is not currently available for this article.
Provided by the Springer Nature SharedIt content-sharing initiative
Biological Research
ISSN: 0717-6287
- Submission enquiries: Access here and click Contact Us
- General enquiries: [email protected]
- Open access
- Published: 27 May 2024
Current status of community resources and priorities for weed genomics research
- Jacob Montgomery 1 ,
- Sarah Morran 1 ,
- Dana R. MacGregor ORCID: orcid.org/0000-0003-0543-0408 2 ,
- J. Scott McElroy ORCID: orcid.org/0000-0003-0331-3697 3 ,
- Paul Neve ORCID: orcid.org/0000-0002-3136-5286 4 ,
- Célia Neto ORCID: orcid.org/0000-0003-3256-5228 4 ,
- Martin M. Vila-Aiub ORCID: orcid.org/0000-0003-2118-290X 5 ,
- Maria Victoria Sandoval 5 ,
- Analia I. Menéndez ORCID: orcid.org/0000-0002-9681-0280 6 ,
- Julia M. Kreiner ORCID: orcid.org/0000-0002-8593-1394 7 ,
- Longjiang Fan ORCID: orcid.org/0000-0003-4846-0500 8 ,
- Ana L. Caicedo ORCID: orcid.org/0000-0002-0378-6374 9 ,
- Peter J. Maughan 10 ,
- Bianca Assis Barbosa Martins 11 ,
- Jagoda Mika 11 ,
- Alberto Collavo 11 ,
- Aldo Merotto Jr. ORCID: orcid.org/0000-0002-1581-0669 12 ,
- Nithya K. Subramanian ORCID: orcid.org/0000-0002-1659-7396 13 ,
- Muthukumar V. Bagavathiannan ORCID: orcid.org/0000-0002-1107-7148 13 ,
- Luan Cutti ORCID: orcid.org/0000-0002-2867-7158 14 ,
- Md. Mazharul Islam 15 ,
- Bikram S. Gill ORCID: orcid.org/0000-0003-4510-9459 16 ,
- Robert Cicchillo 17 ,
- Roger Gast 17 ,
- Neeta Soni ORCID: orcid.org/0000-0002-4647-8355 17 ,
- Terry R. Wright ORCID: orcid.org/0000-0002-3969-2812 18 ,
- Gina Zastrow-Hayes 18 ,
- Gregory May 18 ,
- Jenna M. Malone ORCID: orcid.org/0000-0002-9637-2073 19 ,
- Deepmala Sehgal ORCID: orcid.org/0000-0002-4141-1784 20 ,
- Shiv Shankhar Kaundun ORCID: orcid.org/0000-0002-7249-2046 20 ,
- Richard P. Dale 20 ,
- Barend Juan Vorster ORCID: orcid.org/0000-0003-3518-3508 21 ,
- Bodo Peters 11 ,
- Jens Lerchl ORCID: orcid.org/0000-0002-9633-2653 22 ,
- Patrick J. Tranel ORCID: orcid.org/0000-0003-0666-4564 23 ,
- Roland Beffa ORCID: orcid.org/0000-0003-3109-388X 24 ,
- Alexandre Fournier-Level ORCID: orcid.org/0000-0002-6047-7164 25 ,
- Mithila Jugulam ORCID: orcid.org/0000-0003-2065-9067 15 ,
- Kevin Fengler 18 ,
- Victor Llaca ORCID: orcid.org/0000-0003-4822-2924 18 ,
- Eric L. Patterson ORCID: orcid.org/0000-0001-7111-6287 14 &
- Todd A. Gaines ORCID: orcid.org/0000-0003-1485-7665 1
Genome Biology volume 25 , Article number: 139 ( 2024 ) Cite this article
590 Accesses
7 Altmetric
Metrics details
Weeds are attractive models for basic and applied research due to their impacts on agricultural systems and capacity to swiftly adapt in response to anthropogenic selection pressures. Currently, a lack of genomic information precludes research to elucidate the genetic basis of rapid adaptation for important traits like herbicide resistance and stress tolerance and the effect of evolutionary mechanisms on wild populations. The International Weed Genomics Consortium is a collaborative group of scientists focused on developing genomic resources to impact research into sustainable, effective weed control methods and to provide insights about stress tolerance and adaptation to assist crop breeding.
Each year globally, agricultural producers and landscape managers spend billions of US dollars [ 1 , 2 ] and countless hours attempting to control weedy plants and reduce their adverse effects. These management methods range from low-tech (e.g., pulling plants from the soil by hand) to extremely high-tech (e.g., computer vision-controlled spraying of herbicides). Regardless of technology level, effective control methods serve as strong selection pressures on weedy plants and often result in rapid evolution of weed populations resistant to such methods [ 3 , 4 , 5 , 6 , 7 ]. Thus, humans and weeds have been locked in an arms race, where humans develop new or improved control methods and weeds adapt and evolve to circumvent such methods.
Applying genomics to weed science offers a unique opportunity to study rapid adaptation, epigenetic responses, and examples of evolutionary rescue of diverse weedy species in the face of widespread and powerful selective pressures. Furthermore, lessons learned from these studies may also help to develop more sustainable control methods and to improve crop breeding efforts in the face of our ever-changing climate. While other research fields have used genetics and genomics to uncover the basis of many biological traits [ 8 , 9 , 10 , 11 ] and to understand how ecological factors affect evolution [ 12 , 13 ], the field of weed science has lagged behind in the development of genomic tools essential for such studies [ 14 ]. As research in human and crop genetics pushes into the era of pangenomics (i.e., multiple chromosome scale genome assemblies for a single species [ 15 , 16 ]), publicly available genomic information is still lacking or severely limited for the majority of weed species. Recent reviews of current weed genomes identified 26 [ 17 ] and 32 weed species with sequenced genomes [ 18 ]—many assembled to a sub-chromosome level.
Here, we summarize the current state of weed genomics, highlighting cases where genomics approaches have successfully provided insights on topics such as population genetic dynamics, genome evolution, and the genetic basis of herbicide resistance, rapid adaptation, and crop dedomestication. These highlighted investigations all relied upon genomic resources that are relatively rare for weedy species. Throughout, we identify additional resources that would advance the field of weed science and enable further progress in weed genomics. We then introduce the International Weed Genomics Consortium (IWGC), an open collaboration among researchers, and describe current efforts to generate these additional resources.
Evolution of weediness: potential research utilizing weed genomics tools
Weeds can evolve from non-weed progenitors through wild colonization, crop de-domestication, or crop-wild hybridization [ 19 ]. Because the time span in which weeds have evolved is necessarily limited by the origins of agriculture, these non-weed relatives often still exist and can be leveraged through population genomic and comparative genomic approaches to identify the adaptive changes that have driven the evolution of weediness. The ability to rapidly adapt, persist, and spread in agroecosystems are defining features of weedy plants, leading many to advocate agricultural weeds as ideal candidates for studying rapid plant adaptation [ 20 , 21 , 22 , 23 ]. The insights gained from applying plant ecological approaches to the study of rapid weed adaptation will move us towards the ultimate goals of mitigating such adaptation and increasing the efficacy of crop breeding and biotechnology [ 14 ].
Biology and ecological genomics of weeds
The impressive community effort to create and maintain resources for Arabidopsis thaliana ecological genomics provides a motivating example for the emerging study of weed genomics [ 24 , 25 , 26 , 27 ]. Arabidopsis thaliana was the first flowering plant species to have its genome fully sequenced [ 28 ] and rapidly became a model organism for plant molecular biology. As weedy genomes become available, collection, maintenance, and resequencing of globally distributed accessions of these species will help to replicate the success found in ecological studies of A. thaliana [ 29 , 30 , 31 , 32 , 33 , 34 , 35 ]. Evaluation of these accessions for traits of interest to produce large phenomics data sets (as in [ 36 , 37 , 38 , 39 , 40 ]) enables genome-wide association studies and population genomics analyses aimed at dissecting the genetic basis of variation in such traits [ 41 ]. Increasingly, these resources (e.g. the 1001 genomes project [ 29 ]) have enabled A. thaliana to be utilized as a model species to explore the eco-evolutionary basis of plant adaptation in a more realistic ecological context. Weedy species should supplement lessons in eco-evolutionary genomics learned from these experiments in A. thaliana .
Untargeted genomic approaches for understanding the evolutionary trajectories of populations and the genetic basis of traits as described above rely on the collection of genotypic information from across the genome of many individuals. While whole-genome resequencing accomplishes this requirement and requires no custom methodology, this approach provides more information than is necessary and is prohibitively expensive in species with large genomes. Development and optimization of genotype-by-sequencing methods for capturing reduced representations of newly sequence genomes like those described by [ 42 , 43 , 44 ] will reduce the cost and computational requirements of genetic mapping and population genetic experiments. Most major weed species do not currently have protocols for stable transformation, a key development in the popularity of A. thaliana as a model organism and a requirement for many functional genomic approaches. Functional validation of genes/variants believed to be responsible for traits of interest in weeds has thus far relied on transiently manipulating endogenous gene expression [ 45 , 46 ] or ectopic expression of a transgene in a model system [ 47 , 48 , 49 ]. While these methods have been successful, few weed species have well-studied viral vectors to adapt for use in virus induced gene silencing. Spray induced gene silencing is another potential option for functional investigation of candidate genes in weeds, but more research is needed to establish reliable delivery and gene knockdown [ 50 ]. Furthermore, traits with complex genetic architecture divergent between the researched and model species may not be amenable to functional genomic approaches using transgenesis techniques in model systems. Developing protocols for reduced representation sequencing, stable transformation, and gene editing/silencing in weeds will allow for more thorough characterization of candidate genetic variants underlying traits of interest.
Beyond rapid adaptation, some weedy species offer an opportunity to better understand co-evolution, like that between plants and pollinators and how their interaction leads to the spread of weedy alleles (Additional File 1 : Table S1). A suite of plant–insect traits has co-evolved to maximize the attraction of the insect pollinator community and the efficiency of pollen deposition between flowers ensuring fruit and seed production in many weeds [ 51 , 52 ]. Genetic mapping experiments have identified genes and genetic variants responsible for many floral traits affecting pollinator interaction including petal color [ 53 , 54 , 55 , 56 ], flower symmetry and size [ 57 , 58 , 59 ], and production of volatile organic compounds [ 60 , 61 , 62 ] and nectar [ 63 , 64 , 65 ]. While these studies reveal candidate genes for selection under co-evolution, herbicide resistance alleles may also have pleiotropic effects on the ecology of weeds [ 66 ], altering plant-pollinator interactions [ 67 ]. Discovery of genes and genetic variants involved in weed-pollinator interaction and their molecular and environmental control may create opportunities for better management of weeds with insect-mediated pollination. For example, if management can disrupt pollinator attraction/interaction with these weeds, the efficiency of reproduction may be reduced.
A more complete understanding of weed ecological genomics will undoubtedly elucidate many unresolved questions regarding the genetic basis of various aspects of weediness. For instance, when comparing populations of a species from agricultural and non-agricultural environments, is there evidence for contemporary evolution of weedy traits selected by agricultural management or were “natural” populations pre-adapted to agroecosystems? Where there is differentiation between weedy and natural populations, which traits are under selection and what is the genetic basis of variation in those traits? When comparing between weedy populations, is there evidence for parallel versus non-parallel evolution of weediness at the phenotypic and genotypic levels? Such studies may uncover fundamental truths about weediness. For example, is there a common phenotypic and/or genotypic basis for aspects of weediness among diverse weed species? The availability of characterized accessions and reference genomes for species of interest are required for such studies but only a few weedy species have these resources developed.
Population genomics
Weed species are certainly fierce competitors, able to outcompete crops and endemic species in their native environment, but they are also remarkable colonizers of perturbed habitats. Weeds achieve this through high fecundity, often producing tens of thousands of seeds per individual plant [ 68 , 69 , 70 ]. These large numbers in terms of demographic population size often combine with outcrossing reproduction to generate high levels of diversity with local effective population sizes in the hundreds of thousands [ 71 , 72 ]. This has two important consequences: weed populations retain standing genetic variation and generate many new mutations, supporting weed success in the face of harsh control. The generation of genomic tools to monitor weed populations at the molecular level is a game-changer to understanding weed dynamics and precisely testing the effect of artificial selection (i.e., management) and other evolutionary mechanisms on the genetic make-up of populations.
Population genomic data, without any environmental or phenotypic information, can be used to scan the genomes of weed and non-weed relatives to identify selective sweeps, pointing at loci supporting weed adaptation on micro- or macro-evolutionary scales. Two recent within-species examples include weedy rice, where population differentiation between weedy and domesticated populations was used to identify the genetic basis of weedy de-domestication [ 73 ], and common waterhemp, where consistent allelic differences among natural and agricultural collections resolved a complex set of agriculturally adaptive alleles [ 74 , 75 ]. A recent comparative population genomic study of weedy barnyardgrass and crop millet species has demonstrated how inter-specific investigations can resolve the signatures of crop and weed evolution [ 76 ] (also see [ 77 ] for a non-weed climate adaptation example). Multiple sequence alignments across numerous species provide complementary insight into adaptive convergence over deeper timescales, even with just one genomic sample per species (e.g., [ 78 , 79 ]). Thus, newly sequenced weed genomes combined with genomes available for closely related crops (outlined by [ 14 , 80 ]) and an effort to identify other non-weed wild relatives will be invaluable in characterizing the genetic architecture of weed adaptation and evolution across diverse species.
Weeds experience high levels of genetic selection, both artificial in response to agricultural practices and particularly herbicides, and natural in response to the environmental conditions they encounter [ 81 , 82 ]. Using genomic analysis to identify loci that are the targets of selection, whether natural or artificial, would point at vulnerabilities that could be leveraged against weeds to develop new and more sustainable management strategies [ 83 ]. This is a key motivation to develop genotype-by-environment association (GEA) and selective sweep scan approaches, which allow researchers to resolve the molecular basis of multi-dimensional adaptation [ 84 , 85 ]. GEA approaches, in particular, have been widely used on landscape-wide resequencing collections to determine the genetic basis of climate adaptation (e.g., [ 27 , 86 , 87 ]), but have yet to be fully exploited to diagnose the genetic basis of the various aspects of weediness [ 88 ]. Armed with data on environmental dimensions of agricultural settings, such as focal crop, soil quality, herbicide use, and climate, GEA approaches can help disentangle how discrete farming practices have influenced the evolution of weediness and resolve broader patterns of local adaptation across a weed’s range. Although non-weedy relatives are not technically required for GEA analyses, inclusion of environmental and genomic data from weed progenitors can further distinguish genetic variants underpinning weed origins from those involved in local adaptation.
New weeds emerge frequently [ 89 ], either through hybridization between species as documented for sea beet ( Beta vulgaris ssp. maritima) hybridizing with crop beet to produce progeny that are well adapted to agricultural conditions [ 90 , 91 , 92 ], or through the invasion of alien species that find a new range to colonize. Biosecurity measures are often in place to stop the introduction of new weeds; however, the vast scale of global agricultural commodity trade precludes the possibility of total control. Population genomic analysis is now able to measure gene flow between populations [ 74 , 93 , 94 , 95 ] and identify populations of origin for invasive species including weeds [ 96 , 97 , 98 ]. For example, the invasion route of the pest fruitfly Drosophila suzukii from Eastern Asia to North America and Europe through Hawaii was deciphered using Approximate Bayesian Computation on high-throughput sequencing data from a global sample of multiple populations [ 99 ]. Genomics can also be leveraged to predict invasion rather than explain it. The resequencing of a global sample of common ragweed ( Ambrosia artemisiifolia L.) elucidated a complex invasion route whereby Europe was invaded by multiple introductions of American ragweed that hybridized in Europe prior to a subsequent introduction to Australia [ 100 , 101 ]. In this context, the use of genomically informed species distribution models helps assess the risk associated with different source populations, which in the case of common ragweed, suggests that a source population from Florida would allow ragweed to invade most of northern Australia [ 102 ]. Globally coordinated research efforts to understand potential distribution models could support the transformation of biosecurity from perspective analysis towards predictive risk assessment.
Herbicide resistance and weed management
Herbicide resistance is among the numerous weedy traits that can evolve in plant populations exposed to agricultural selection pressures. Over-reliance on herbicides to control weeds, along with low diversity and lack of redundancy in weed management strategies, has resulted in globally widespread herbicide resistance [ 103 ]. To date, 272 herbicide-resistant weed species have been reported worldwide, and at least one resistance case exists for 21 of the 31 existing herbicide sites of action [ 104 ]—significantly limiting chemical weed control options available to agriculturalists. This limitation of control options is exacerbated by the recent lack of discovery of herbicides with new sites of action [ 105 ].
Herbicide resistance may result from several different physiological mechanisms. Such mechanisms have been classified into two main groups, target-site resistance (TSR) [ 4 , 106 ] and non-target-site resistance (NTSR) [ 4 , 107 ]. The first group encompasses changes that reduce binding affinity between a herbicide and its target [ 108 ]. These changes may provide resistance to multiple herbicides that have a common biochemical target [ 109 ] and can be effectively managed through mixture and/or rotation of herbicides targeting different sites of action [ 110 ]. The second group (NTSR), includes alterations in herbicide absorption, translocation, sequestration, and/or metabolism that may lead to unpredictable pleotropic cross-resistance profiles where structurally and functionally diverse herbicides are rendered ineffective by one or more genetic variant(s) [ 47 ]. This mechanism of resistance threatens not only the efficacy of existing herbicidal chemistries, but also ones yet to be discovered. While TSR is well understood because of the ease of identification and molecular characterization of target site variants, NTSR mechanisms are significantly more challenging to research because they are often polygenic, and the resistance causing element(s) are not well understood [ 111 ].
Improving the current understanding of metabolic NTSR mechanisms is not an easy task, since genes of diverse biochemical functions are involved, many of which exist as extensive gene families [ 109 , 112 ]. Expression changes of NTSR genes have been implicated in several resistance cases where the protein products of the genes are functionally equivalent across sensitive and resistant plants, but their relative abundance leads to resistance. Thus, regulatory elements of NTSR genes have been scrutinized to understand their role in NTSR mechanisms [ 113 ]. Similarly, epigenetic modifications have been hypothesized to play a role in NTSR, with much remaining to be explored [ 114 , 115 , 116 ]. Untargeted approaches such as genome-wide association, selective sweep scans, linkage mapping, RNA-sequencing, and metabolomic profiling have proven helpful to complement more specific biochemical- and chemo-characterization studies towards the elucidation of NTSR mechanisms as well as their regulation and evolution [ 47 , 117 , 118 , 119 , 120 , 121 , 122 , 123 , 124 ]. Even in cases where resistance has been attributed to TSR, genetic mapping approaches can detect other NTSR loci contributing to resistance (as shown by [ 123 ]) and provide further evidence for the role of TSR mutations across populations. Knowledge of the genetic basis of NTSR will aid the rational design of herbicides by screening new compounds for interaction with newly discovered NTSR proteins during early research phases and by identifying conserved chemical structures that interact with these proteins that should be avoided in small molecule design.
Genomic resources can also be used to predict the protein structure for novel herbicide target site and metabolism genes. This will allow for prediction of efficacy and selectivity for new candidate herbicides in silico to increase herbicide discovery throughput as well as aid in the design and development of next-generation technologies for sustainable weed management. Proteolysis targeting chimeras (PROTACs) have the potential to bind desired targets with great selectivity and degrade proteins by utilizing natural protein ubiquitination and degradation pathways within plants [ 125 ]. Spray-induced gene silencing in weeds using oligonucleotides has potential as a new, innovative, and sustainable method for weed management, but improved methods for design and delivery of oligonucleotides are needed to make this technique a viable management option [ 50 ]. Additionally, success in the field of pharmaceutical drug discovery in the development of molecules modulating protein–protein interactions offers another potential avenue towards the development of herbicides with novel targets [ 126 , 127 ]. High-quality reference genomes allow for the design of new weed management technologies like the ones listed here that are specific to—and effective across—weed species but have a null effect on non-target organisms.
Comparative genomics and genome biology
The genomes of weed species are as diverse as weed species themselves. Weeds are found across highly diverged plant families and often have no phylogenetically close model or crop species relatives for comparison. On all measurable metrics, weed genomes run the gamut. Some have smaller genomes like Cyperus spp. (~ 0.26 Gb) while others are larger, such as Avena fatua (~ 11.1 Gb) (Table 1 ). Some have high heterozygosity in terms of single-nucleotide polymorphisms, such as the Amaranthus spp., while others are primarily self-pollinated and quite homozygous, such as Poa annua [ 128 , 129 ]. Some are diploid such as Conyza canadensis and Echinochloa haploclada while others are polyploid such as C. sumetrensis , E. crus-galli , and E. colona [ 76 ]. The availability of genomic resources in these diverse, unexplored branches of the tree of life allows us to identify consistencies and anomalies in the field of genome biology.
The weed genomes published so far have focused mainly on weeds of agronomic crops, and studies have revolved around their ability to resist key herbicides. For example, genomic resources were vital in the elucidation of herbicide resistance cases involving target site gene copy number variants (CNVs). Gene CNVs of 5-enolpyruvylshikimate-3-phosphate synthase ( EPSPS ) have been found to confer resistance to the herbicide glyphosate in diverse weed species. To date, nine species have independently evolved EPSPS CNVs, and species achieve increased EPSPS copy number via different mechanisms [ 153 ]. For instance, the EPSPS CNV in Bassia scoparia is caused by tandem duplication, which is accredited to transposable element insertions flanking EPSPS and subsequent unequal crossing over events [ 154 , 155 ]. In Eleusine indica , a EPSPS CNV was caused by translocation of the EPSPS locus into the subtelomere followed by telomeric sequence exchange [ 156 ]. One of the most fascinating genome biology discoveries in weed science has been that of extra-chromosomal circular DNAs (eccDNAs) that harbor the EPSPS gene in the weed species Amaranthus palmeri [ 157 , 158 ]. In this case, the eccDNAs autonomously replicate separately from the nuclear genome and do not reintegrate into chromosomes, which has implications for inheritance, fitness, and genome structure [ 159 ]. These discoveries would not have been possible without reference assemblies of weed genomes, next-generation sequencing, and collaboration with experts in plant genomics and bioinformatics.
Another question that is often explored with weedy genomes is the nature and composition of gene families that are associated with NTSR. Gene families under consideration often include cytochrome P450s (CYPs), glutathione- S -transferases (GSTs), ABC transporters, etc. Some questions commonly considered with new weed genomes include how many genes are in each of these gene families, where are they located, and which weed accessions and species have an over-abundance of them that might explain their ability to evolve resistance so rapidly [ 76 , 146 , 160 , 161 ]? Weed genome resources are necessary to answer questions about gene family expansion or contraction during the evolution of weediness, including the role of polyploidy in NTSR gene family expansion as explored by [ 162 ].
Translational research and communication with weed management stakeholders
Whereas genomics of model plants is typically aimed at addressing fundamental questions in plant biology, and genomics of crop species has the obvious goal of crop improvement, goals of genomics of weedy plants also include the development of more effective and sustainable strategies for their management. Weed genomic resources assist with these objectives by providing novel molecular ecological and evolutionary insights from the context of intensive anthropogenic management (which is lacking in model plants), and offer knowledge and resources for trait discovery for crop improvement, especially given that many wild crop relatives are also important agronomic weeds (e.g., [ 163 ]). For instance, crop-wild relatives are valuable for improving crop breeding for marginal environments [ 164 ]. Thus, weed genomics presents unique opportunities and challenges relative to plant genomics more broadly. It should also be noted that although weed science at its core is an applied discipline, it draws broadly from many scientific disciplines such as, plant physiology, chemistry, ecology, and evolutionary biology, to name a few. The successful integration of weed-management strategies, therefore, requires extensive collaboration among individuals collectively possessing the necessary expertise [ 165 ].
With the growing complexity of herbicide resistance management, practitioners are beginning to recognize the importance of understanding resistance mechanisms to inform appropriate management tactics [ 14 ]. Although weed science practitioners do not need to understand the technical details of weed genomics, their appreciation of the power of weed genomics—together with their unique insights from field observations—will yield novel opportunities for applications of weed genomics to weed management. In particular, combining field management history with information on weed resistance mechanisms is expected to provide novel insights into evolutionary trajectories (e.g. [ 6 , 166 ]), which can be utilized for disrupting evolutionary adaptation. It can be difficult to obtain field history information from practitioners, but developing an understanding among them of the importance of such information can be invaluable.
Development of weed genomics resources by the IWGC
Weed genomics is a fast-growing field of research with many recent breakthroughs and many unexplored areas of study. The International Weed Genomics Consortium (IWGC) started in 2021 to address the roadblocks listed above and to promote the study of weedy plants. The IWGC is an open collaboration among academic, government, and industry researchers focused on producing genomic tools for weedy species from around the world. Through this collaboration, our initial aim is to provide chromosome-level reference genome assemblies for at least 50 important weedy species from across the globe that are chosen based on member input, economic impact, and global prevalence (Fig. 1 ). Each genome will include annotation of gene models and repetitive elements and will be freely available through public databases with no intellectual property restrictions. Additionally, future funding of the IWGC will focus on improving gene annotations and supplementing these reference genomes with tools that increase their utility.

The International Weed Genomics Consortium (IWGC) collected input from the weed genomics community to develop plans for weed genome sequencing, annotation, user-friendly genome analysis tools, and community engagement
Reference genomes and data analysis tools
The first objective of the IWGC is to provide high-quality genomic resources for agriculturally important weeds. The IWGC therefore created two main resources for information about, access to, or analysis of weed genomic data (Fig. 1 ). The IWGC website (available at [ 167 ]) communicates the status and results of genome sequencing projects, information on training and funding opportunities, upcoming events, and news in weed genomics. It also contains details of all sequenced species including genome size, ploidy, chromosome number, herbicide resistance status, and reference genome assembly statistics. The IWGC either compiles existing data on genome size, ploidy, and chromosome number, or obtains the data using flow cytometry and cytogenetics (Fig. 1 ; Additional File 2 : Fig S1-S4). Through this website, users can request an account to access our second main resource, an online genome database called WeedPedia (accessible at [ 168 ]), with an account that is created within 3–5 working days of an account request submission. WeedPedia hosts IWGC-generated and other relevant publicly accessible genomic data as well as a suite of bioinformatic tools. Unlike what is available for other fields, weed science did not have a centralized hub for genomics information, data, and analysis prior to the IWGC. Our intention in creating WeedPedia is to encourage collaboration and equity of access to information across the research community. Importantly, all genome assemblies and annotations from the IWGC (Table 1 ), along with the raw data used to produce them, will be made available through NCBI GenBank. Upon completion of a 1-year sponsoring member data confidentiality period for each species (dates listed in Table 1 ), scientific teams within the IWGC produce the first genome-wide investigation to submit for publication including whole genome level analyses on genes, gene families, and repetitive sequences as well as comparative analysis with other species. Genome assemblies and data will be publicly available through NCBI as part of these initial publications for each species.
WeedPedia is a cloud-based omics database management platform built from the software “CropPedia” and licensed from KeyGene (Wageningen, The Netherlands). The interface allows users to access, visualize, and download genome assemblies along with structural and functional annotation. The platform includes a genome browser, comparative map viewer, pangenome tools, RNA-sequencing data visualization tools, genetic mapping and marker analysis tools, and alignment capabilities that allow searches by keyword or sequence. Additionally, genes encoding known target sites of herbicides have been specially annotated, allowing users to quickly identify and compare these genes of interest. The platform is flexible, making it compatible with future integration of other data types such as epigenetic or proteomic information. As an online platform with a graphical user interface, WeedPedia provides user-friendly, intuitive tools that encourage users to integrate genomics into their research while also allowing more advanced users to download genomic data to be used in custom analysis pipelines. We aspire for WeedPedia to mimic the success of other public genomic databases such as NCBI, CoGe, Phytozome, InsectBase, and Mycocosm to name a few. WeedPedia currently hosts reference genomes for 40 species (some of which are currently in their 1-year confidentiality period) with additional genomes in the pipeline to reach a currently planned total of 55 species (Table 1 ). These genomes include both de novo reference genomes generated or in progress by the IWGC (31 species; Table 1 ), and publicly available genome assemblies of 24 weedy or related species that were generated by independent research groups (Table 2 ). As of May 2024, WeedPedia has over 370 registered users from more than 27 countries spread across 6 continents.
The IWGC reference genomes are generated in partnership with the Corteva Agriscience Genome Center of Excellence (Johnston, Iowa) using a combination of single-molecule long-read sequencing, optical genome maps, and chromosome conformation mapping. This strategy has already yielded highly contiguous, phased, chromosome-level assemblies for 26 weed species, with additional assemblies currently in progress (Table 1 ). The IWGC assemblies have been completed as single or haplotype-resolved double-haplotype pseudomolecules in inbreeding and outbreeding species, respectively, with multiple genomes being near gapless. For example, the de novo assemblies of the allohexaploids Conyza sumatrensis and Chenopodium album have all chromosomes captured in single scaffolds and most chromosomes being gapless from telomere to telomere. Complementary full-length isoform (IsoSeq) sequencing of RNA collected from diverse tissue types and developmental stages assists in the development of gene models during annotation.
As with accessibility of data, a core objective of the IWGC is to facilitate open access to sequenced germplasm when possible for featured species. Historically, the weed science community has rarely shared or adopted standard germplasm (e.g., specific weed accessions). The IWGC has selected a specific accession of each species for reference genome assembly (typically susceptible to herbicides). In collaboration with a parallel effort by the Herbicide Resistant Plants committee of the Weed Science Society of America, seeds of the sequenced weed accessions will be deposited in the United States Department of Agriculture Germplasm Resources Information Network [ 186 ] for broad access by the scientific community and their accession numbers will be listed on the IWGC website. In some cases, it is not possible to generate enough seed to deposit into a public repository (e.g., plants that typically reproduce vegetatively, that are self-incompatible, or that produce very few seeds from a single individual). In these cases, the location of collection for sequenced accessions will at least inform the community where the sequenced individual came from and where they may expect to collect individuals with similar genotypes. The IWGC ensures that sequenced accessions are collected and documented to comply with the Nagoya Protocol on access to genetic resources and the fair and equitable sharing of benefits arising from their utilization under the Convention on Biological Diversity and related Access and Benefit Sharing Legislation [ 187 ]. As additional accessions of weed species are sequenced (e.g., pangenomes are obtained), the IWGC will facilitate germplasm sharing protocols to support collaboration. Further, to simplify the investigation of herbicide resistance, the IWGC will link WeedPedia with the International Herbicide-Resistant Weed Database [ 104 ], an already widely known and utilized database for weed scientists.
Training and collaboration in weed genomics
Beyond producing genomic tools and resources, a priority of the IWGC is to enable the utilization of these resources across a wide range of stakeholders. A holistic approach to training is required for weed science generally [ 188 ], and we would argue even more so for weed genomics. To accomplish our training goals, the IWGC is developing and delivering programs aimed at the full range of IWGC stakeholders and covering a breadth of relevant topics. We have taken care to ensure our approaches are diverse as to provide training to researchers with all levels of existing experience and differing reasons for engaging with these tools. Throughout, the focus is on ensuring that our training and outreach result in impacts that benefit a wide range of stakeholders.
Although recently developed tools are incredibly enabling and have great potential to replace antiquated methodology [ 189 ] and to solve pressing weed science problems [ 14 ], specialized computational skills are required to fully explore and unlock meaning from these highly complex datasets. Collaboration with, or training of, computational biologists equipped with these skills and resources developed by the IWGC will enable weed scientists to expand research programs and better understand the genetic underpinnings of weed evolution and herbicide resistance. To fill existing skill gaps, the IWGC is developing summer bootcamps and online modules directed specifically at weed scientists that will provide training on computational skills (Fig. 1 ). Because successful utilization of the IWGC resources requires more than general computational skills, we have created three targeted workshops that teach practical skills related to genomics databases, molecular biology, and population genomics (available at [ 190 ]). The IWGC has also hosted two official conference meetings, one in September of 2021 and one in January of 2023, with more conferences planned. These conferences have included invited speakers to present successful implementations of weed genomics, educational workshops to build computational skills, and networking opportunities for research to connect and collaborate.
Engagement opportunities during undergraduate degrees have been shown to improve academic outcomes [ 191 , 192 ]. As one activity to help achieve this goal, the IWGC has sponsored opportunities for US undergraduates to undertake a 10-week research experience, which includes an introduction to bioinformatics, a plant genomics research project that results in a presentation, and access to career building opportunities in diverse workplace environments. To increase equitable access to conferences and professional communities, we supported early career researchers to attend the first two IWGC conferences in the USA as well as workshops and bootcamps in Europe, South America, and Australia. These hybrid or in-person travel grants are intentionally designed to remove barriers and increase participation of individuals from backgrounds and experiences currently underrepresented within weed/plant science or genomics [ 193 ]. Recipients of these travel awards gave presentations and gained the measurable benefits that come from either virtual or in-person participation in conferences [ 194 ]. Moving forward, weed scientists must amass skills associated with genomic analyses and collaborate with other area experts to fully leverage resources developed by the IWGC.
The tools generated through the IWGC will enable many new research projects with diverse objectives like those listed above. In summary, contiguous genome assemblies and complete annotation information will allow weed scientists to join plant breeders in the use of genetic mapping for many traits including stress tolerance, plant architecture, and herbicide resistance (especially important for cases of NTSR). These assemblies will also allow for investigations of population structure, gene flow, and responses to evolutionary mechanisms like genetic bottlenecking and artificial selection. Understanding gene sequences across diverse weed species will be vital in modeling new herbicide target site proteins and designing novel effective herbicides with minimal off-target effects. The IWGC website will improve accessibility to weed genomics data by providing a single hub for reference genomes as well as phenotypic and genotypic information for accessions shared with the IWGC. Deposition of sequenced germplasm into public repositories will ensure that researchers are able to access and utilize these accessions in their own research to make the field more standardized and equitable. WeedPedia allows users of all backgrounds to quickly access information of interest such as herbicide target site gene sequence or subcellular localization of protein products for different genes. Users can also utilize server-based tools such as BLAST and genome browsing similar to other public genomic databases. Finally, the IWGC is committed to training and connecting weed genomicists through hosting trainings, workshops, and conferences.
Conclusions
Weeds are unique and fascinating plants, having significant impacts on agriculture and ecosystems; and yet, aspects of their biology, ecology, and genetics remain poorly understood. Weeds represent a unique area within plant biology, given their repeated rapid adaptation to sudden and severe shifts in the selective landscape of anthropogenic management practices. The production of a public genomics database with reference genomes and annotations for over 50 weed species represents a substantial step forward towards research goals that improve our understanding of the biology and evolution of weeds. Future work is needed to improve annotations, particularly for complex gene families involved in herbicide detoxification, structural variants, and mobile genetic elements. As reference genome assemblies become available; standard, affordable methods for gathering genotype information will allow for the identification of genetic variants underlying traits of interest. Further, methods for functional validation and hypothesis testing are needed in weeds to validate the effect of genetic variants detected through such experiments, including systems for transformation, gene editing, and transient gene silencing and expression. Future research should focus on utilizing weed genomes to investigate questions about evolutionary biology, ecology, genetics of weedy traits, and weed population dynamics. The IWGC plans to continue the public–private partnership model to host the WeedPedia database over time, integrate new datasets such as genome resequencing and transcriptomes, conduct trainings, and serve as a research coordination network to ensure that advances in weed science from around the world are shared across the research community (Fig. 1 ). Bridging basic plant genomics with translational applications in weeds is needed to deliver on the potential of weed genomics to improve weed management and crop breeding.
Availability of data and materials
All genome assemblies and related sequencing data produced by the IWGC will be available through NCBI as part of publications reporting the first genome-wide analysis for each species.
Gianessi LP, Nathan PR. The value of herbicides in U.S. crop production. Weed Technol. 2007;21(2):559–66.
Article Google Scholar
Pimentel D, Lach L, Zuniga R, Morrison D. Environmental and economic costs of nonindigenous species in the United States. Bioscience. 2000;50(1):53–65.
Barrett SH. Crop mimicry in weeds. Econ Bot. 1983;37(3):255–82.
Powles SB, Yu Q. Evolution in action: plants resistant to herbicides. Annu Rev Plant Biol. 2010;61:317–47.
Article CAS PubMed Google Scholar
Thurber CS, Reagon M, Gross BL, Olsen KM, Jia Y, Caicedo AL. Molecular evolution of shattering loci in U.S. weedy rice. Mol Ecol. 2010;19(16):3271–84.
Article PubMed PubMed Central Google Scholar
Comont D, Lowe C, Hull R, Crook L, Hicks HL, Onkokesung N, et al. Evolution of generalist resistance to herbicide mixtures reveals a trade-off in resistance management. Nat Commun. 2020;11(1):3086.
Article CAS PubMed PubMed Central Google Scholar
Ashworth MB, Walsh MJ, Flower KC, Vila-Aiub MM, Powles SB. Directional selection for flowering time leads to adaptive evolution in Raphanus raphanistrum (wild radish). Evol Appl. 2016;9(4):619–29.
Chan EK, Rowe HC, Kliebenstein DJ. Understanding the evolution of defense metabolites in Arabidopsis thaliana using genome-wide association mapping. Genetics. 2010;185(3):991–1007.
Frayling TM, Timpson NJ, Weedon MN, Zeggini E, Freathy RM, Lindgren CM, et al. A common variant in the FTO gene is associated with body mass index and predisposes to childhood and adult obesity. Science. 2007;316(5826):889–94.
Harkess A, Zhou J, Xu C, Bowers JE, Van der Hulst R, Ayyampalayam S, et al. The asparagus genome sheds light on the origin and evolution of a young Y chromosome. Nat Commun. 2017;8(1):1279.
Periyannan S, Moore J, Ayliffe M, Bansal U, Wang X, Huang L, et al. The gene Sr33 , an ortholog of barley Mla genes, encodes resistance to wheat stem rust race Ug99. Science. 2013;341(6147):786–8.
Ågren J, Oakley CG, McKay JK, Lovell JT, Schemske DW. Genetic mapping of adaptation reveals fitness tradeoffs in Arabidopsis thaliana . Proc Natl Acad Sci U S A. 2013;110(52):21077–82.
Article PubMed Central Google Scholar
Schartl M, Walter RB, Shen Y, Garcia T, Catchen J, Amores A, et al. The genome of the platyfish, Xiphophorus maculatus , provides insights into evolutionary adaptation and several complex traits. Nat Genet. 2013;45(5):567–72.
Ravet K, Patterson EL, Krähmer H, Hamouzová K, Fan L, Jasieniuk M, et al. The power and potential of genomics in weed biology and management. Pest Manag Sci. 2018;74(10):2216–25.
Hufford MB, Seetharam AS, Woodhouse MR, Chougule KM, Ou S, Liu J, et al. De novo assembly, annotation, and comparative analysis of 26 diverse maize genomes. Science. 2021;373(6555):655–62.
Liao W-W, Asri M, Ebler J, Doerr D, Haukness M, Hickey G, et al. A draft human pangenome reference. Nature. 2023;617(7960):312–24.
Huang Y, Wu D, Huang Z, Li X, Merotto A, Bai L, et al. Weed genomics: yielding insights into the genetics of weedy traits for crop improvement. aBIOTECH. 2023;4:20–30.
Chen K, Yang H, Wu D, Peng Y, Lian L, Bai L, et al. Weed biology and management in the multi-omics era: progress and perspectives. Plant Commun. 2024;5(4):100816.
De Wet JMJ, Harlan JR. Weeds and domesticates: evolution in the man-made habitat. Econ Bot. 1975;29(2):99–108.
Mahaut L, Cheptou PO, Fried G, Munoz F, Storkey J, Vasseur F, et al. Weeds: against the rules? Trends Plant Sci. 2020;25(11):1107–16.
Neve P, Vila-Aiub M, Roux F. Evolutionary-thinking in agricultural weed management. New Phytol. 2009;184(4):783–93.
Article PubMed Google Scholar
Sharma G, Barney JN, Westwood JH, Haak DC. Into the weeds: new insights in plant stress. Trends Plant Sci. 2021;26(10):1050–60.
Vigueira CC, Olsen KM, Caicedo AL. The red queen in the corn: agricultural weeds as models of rapid adaptive evolution. Heredity (Edinb). 2013;110(4):303–11.
Donohue K, Dorn L, Griffith C, Kim E, Aguilera A, Polisetty CR, et al. Niche construction through germination cueing: life-history responses to timing of germination in Arabidopsis thaliana . Evolution. 2005;59(4):771–85.
PubMed Google Scholar
Exposito-Alonso M. Seasonal timing adaptation across the geographic range of Arabidopsis thaliana . Proc Natl Acad Sci U S A. 2020;117(18):9665–7.
Fournier-Level A, Korte A, Cooper MD, Nordborg M, Schmitt J, Wilczek AM. A map of local adaptation in Arabidopsis thaliana . Science. 2011;334(6052):86–9.
Hancock AM, Brachi B, Faure N, Horton MW, Jarymowycz LB, Sperone FG, et al. Adaptation to climate across the Arabidopsis thaliana genome. Science. 2011;334(6052):83–6.
Initiative TAG. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana . Nature. 2000;408(6814):796–815.
Alonso-Blanco C, Andrade J, Becker C, Bemm F, Bergelson J, Borgwardt KM, et al. 1,135 genomes reveal the global pattern of polymorphism in Arabidopsis thaliana . Cell. 2016;166(2):481–91.
Durvasula A, Fulgione A, Gutaker RM, Alacakaptan SI, Flood PJ, Neto C, et al. African genomes illuminate the early history and transition to selfing in Arabidopsis thaliana . Proc Natl Acad Sci U S A. 2017;114(20):5213–8.
Frachon L, Mayjonade B, Bartoli C, Hautekèete N-C, Roux F. Adaptation to plant communities across the genome of Arabidopsis thaliana . Mol Biol Evol. 2019;36(7):1442–56.
Fulgione A, Koornneef M, Roux F, Hermisson J, Hancock AM. Madeiran Arabidopsis thaliana reveals ancient long-range colonization and clarifies demography in Eurasia. Mol Biol Evol. 2018;35(3):564–74.
Fulgione A, Neto C, Elfarargi AF, Tergemina E, Ansari S, Göktay M, et al. Parallel reduction in flowering time from de novo mutations enable evolutionary rescue in colonizing lineages. Nat Commun. 2022;13(1):1461.
Kasulin L, Rowan BA, León RJC, Schuenemann VJ, Weigel D, Botto JF. A single haplotype hyposensitive to light and requiring strong vernalization dominates Arabidopsis thaliana populations in Patagonia. Argentina Mol Ecol. 2017;26(13):3389–404.
Picó FX, Méndez-Vigo B, Martínez-Zapater JM, Alonso-Blanco C. Natural genetic variation of Arabidopsis thaliana is geographically structured in the Iberian peninsula. Genetics. 2008;180(2):1009–21.
Atwell S, Huang YS, Vilhjálmsson BJ, Willems G, Horton M, Li Y, et al. Genome-wide association study of 107 phenotypes in Arabidopsis thaliana inbred lines. Nature. 2010;465(7298):627–31.
Flood PJ, Kruijer W, Schnabel SK, van der Schoor R, Jalink H, Snel JFH, et al. Phenomics for photosynthesis, growth and reflectance in Arabidopsis thaliana reveals circadian and long-term fluctuations in heritability. Plant Methods. 2016;12(1):14.
Marchadier E, Hanemian M, Tisné S, Bach L, Bazakos C, Gilbault E, et al. The complex genetic architecture of shoot growth natural variation in Arabidopsis thaliana . PLoS Genet. 2019;15(4):e1007954.
Tisné S, Serrand Y, Bach L, Gilbault E, Ben Ameur R, Balasse H, et al. Phenoscope: an automated large-scale phenotyping platform offering high spatial homogeneity. Plant J. 2013;74(3):534–44.
Tschiersch H, Junker A, Meyer RC, Altmann T. Establishment of integrated protocols for automated high throughput kinetic chlorophyll fluorescence analyses. Plant Methods. 2017;13:54.
Chen X, MacGregor DR, Stefanato FL, Zhang N, Barros-Galvão T, Penfield S. A VEL3 histone deacetylase complex establishes a maternal epigenetic state controlling progeny seed dormancy. Nat Commun. 2023;14(1):2220.
Choi M, Scholl UI, Ji W, Liu T, Tikhonova IR, Zumbo P, et al. Genetic diagnosis by whole exome capture and massively parallel DNA sequencing. Proc Natl Acad Sci U S A. 2009;106(45):19096–101.
Davey JW, Blaxter ML. RADSeq: next-generation population genetics. Brief Funct Genomics. 2010;9(5–6):416–23.
Elshire RJ, Glaubitz JC, Sun Q, Poland JA, Kawamoto K, Buckler ES, et al. A robust, simple genotyping-by-sequencing (GBS) approach for high diversity species. PLoS ONE. 2011;6(5):e19379.
MacGregor DR. What makes a weed a weed? How virus-mediated reverse genetics can help to explore the genetics of weediness. Outlooks Pest Manag. 2020;31(5):224–9.
Mellado-Sánchez M, McDiarmid F, Cardoso V, Kanyuka K, MacGregor DR. Virus-mediated transient expression techniques enable gene function studies in blackgrass. Plant Physiol. 2020;183(2):455–9.
Dimaano NG, Yamaguchi T, Fukunishi K, Tominaga T, Iwakami S. Functional characterization of Cytochrome P450 CYP81A subfamily to disclose the pattern of cross-resistance in Echinochloa phyllopogon . Plant Mol Biol. 2020;102(4–5):403–16.
de Figueiredo MRA, Küpper A, Malone JM, Petrovic T, de Figueiredo ABTB, Campagnola G, et al. An in-frame deletion mutation in the degron tail of auxin coreceptor IAA2 confers resistance to the herbicide 2,4-D in Sisymbrium orientale . Proc Natl Acad Sci U S A. 2022;119(9):e2105819119.
Patzoldt WL, Hager AG, McCormick JS, Tranel PJ. A codon deletion confers resistance to herbicides inhibiting protoporphyrinogen oxidase. Proc Natl Acad Sci U S A. 2006;103(33):12329–34.
Zabala-Pardo D, Gaines T, Lamego FP, Avila LA. RNAi as a tool for weed management: challenges and opportunities. Adv Weed Sci. 2022;40(spe1):e020220096.
Fattorini R, Glover BJ. Molecular mechanisms of pollination biology. Annu Rev Plant Biol. 2020;71:487–515.
Rollin O, Benelli G, Benvenuti S, Decourtye A, Wratten SD, Canale A, et al. Weed-insect pollinator networks as bio-indicators of ecological sustainability in agriculture. A review Agron Sustain Dev. 2016;36(1):8.
Irwin RE, Strauss SY. Flower color microevolution in wild radish: evolutionary response to pollinator-mediated selection. Am Nat. 2005;165(2):225–37.
Ma B, Wu J, Shi T-L, Yang Y-Y, Wang W-B, Zheng Y, et al. Lilac ( Syringa oblata ) genome provides insights into its evolution and molecular mechanism of petal color change. Commun Biol. 2022;5(1):686.
Xing A, Wang X, Nazir MF, Zhang X, Wang X, Yang R, et al. Transcriptomic and metabolomic profiling of flavonoid biosynthesis provides novel insights into petals coloration in Asian cotton ( Gossypium arboreum L.). BMC Plant Biol. 2022;22(1):416.
Zheng Y, Chen Y, Liu Z, Wu H, Jiao F, Xin H, et al. Important roles of key genes and transcription factors in flower color differences of Nicotiana alata . Genes (Basel). 2021;12(12):1976.
Krizek BA, Anderson JT. Control of flower size. J Exp Bot. 2013;64(6):1427–37.
Powell AE, Lenhard M. Control of organ size in plants. Curr Biol. 2012;22(9):R360–7.
Spencer V, Kim M. Re"CYC"ling molecular regulators in the evolution and development of flower symmetry. Semin Cell Dev Biol. 2018;79:16–26.
Amrad A, Moser M, Mandel T, de Vries M, Schuurink RC, Freitas L, et al. Gain and loss of floral scent production through changes in structural genes during pollinator-mediated speciation. Curr Biol. 2016;26(24):3303–12.
Delle-Vedove R, Schatz B, Dufay M. Understanding intraspecific variation of floral scent in light of evolutionary ecology. Ann Bot. 2017;120(1):1–20.
Pichersky E, Gershenzon J. The formation and function of plant volatiles: perfumes for pollinator attraction and defense. Curr Opin Plant Biol. 2002;5(3):237–43.
Ballerini ES, Kramer EM, Hodges SA. Comparative transcriptomics of early petal development across four diverse species of Aquilegia reveal few genes consistently associated with nectar spur development. BMC Genom. 2019;20(1):668.
Corbet SA, Willmer PG, Beament JWL, Unwin DM, Prys-Jones OE. Post-secretory determinants of sugar concentration in nectar. Plant Cell Environ. 1979;2(4):293–308.
Galliot C, Hoballah ME, Kuhlemeier C, Stuurman J. Genetics of flower size and nectar volume in Petunia pollination syndromes. Planta. 2006;225(1):203–12.
Vila-Aiub MM, Neve P, Powles SB. Fitness costs associated with evolved herbicide resistance alleles in plants. New Phytol. 2009;184(4):751–67.
Baucom RS. Evolutionary and ecological insights from herbicide-resistant weeds: what have we learned about plant adaptation, and what is left to uncover? New Phytol. 2019;223(1):68–82.
Bajwa AA, Latif S, Borger C, Iqbal N, Asaduzzaman M, Wu H, et al. The remarkable journey of a weed: biology and management of annual ryegrass ( Lolium rigidum ) in conservation cropping systems of Australia. Plants (Basel). 2021;10(8):1505.
Bitarafan Z, Andreasen C. Fecundity allocation in some european weed species competing with crops. Agronomy. 2022;12(5):1196.
Costea M, Weaver SE, Tardif FJ. The biology of Canadian weeds. 130. Amaranthus retroflexus L., A. powellii , A. powellii S. Watson, and A. hybridus L. Can J Plant Sci. 2004;84(2):631–68.
Dixon A, Comont D, Slavov GT, Neve P. Population genomics of selectively neutral genetic structure and herbicide resistance in UK populations of Alopecurus myosuroides . Pest Manag Sci. 2021;77(3):1520–9.
Kersten S, Chang J, Huber CD, Voichek Y, Lanz C, Hagmaier T, et al. Standing genetic variation fuels rapid evolution of herbicide resistance in blackgrass. Proc Natl Acad Sci U S A. 2023;120(16):e2206808120.
Qiu J, Zhou Y, Mao L, Ye C, Wang W, Zhang J, et al. Genomic variation associated with local adaptation of weedy rice during de-domestication. Nat Commun. 2017;8(1):15323.
Kreiner JM, Caballero A, Wright SI, Stinchcombe JR. Selective ancestral sorting and de novo evolution in the agricultural invasion of Amaranthus tuberculatus . Evolution. 2022;76(1):70–85.
Kreiner JM, Latorre SM, Burbano HA, Stinchcombe JR, Otto SP, Weigel D, et al. Rapid weed adaptation and range expansion in response to agriculture over the past two centuries. Science. 2022;378(6624):1079–85.
Wu D, Shen E, Jiang B, Feng Y, Tang W, Lao S, et al. Genomic insights into the evolution of Echinochloa species as weed and orphan crop. Nat Commun. 2022;13(1):689.
Yeaman S, Hodgins KA, Lotterhos KE, Suren H, Nadeau S, Degner JC, et al. Convergent local adaptation to climate in distantly related conifers. Science. 2016;353(6306):1431–3.
Haudry A, Platts AE, Vello E, Hoen DR, Leclercq M, Williamson RJ, et al. An atlas of over 90,000 conserved noncoding sequences provides insight into crucifer regulatory regions. Nat Genet. 2013;45(8):891–8.
Sackton TB, Grayson P, Cloutier A, Hu Z, Liu JS, Wheeler NE, et al. Convergent regulatory evolution and loss of flight in paleognathous birds. Science. 2019;364(6435):74–8.
Ye CY, Fan L. Orphan crops and their wild relatives in the genomic era. Mol Plant. 2021;14(1):27–39.
Clements DR, Jones VL. Ten ways that weed evolution defies human management efforts amidst a changing climate. Agronomy. 2021;11(2):284.
Article CAS Google Scholar
Weinig C. Rapid evolutionary responses to selection in heterogeneous environments among agricultural and nonagricultural weeds. Int J Plant Sci. 2005;166(4):641–7.
Cousens RD, Fournier-Level A. Herbicide resistance costs: what are we actually measuring and why? Pest Manag Sci. 2018;74(7):1539–46.
Lasky JR, Josephs EB, Morris GP. Genotype–environment associations to reveal the molecular basis of environmental adaptation. Plant Cell. 2023;35(1):125–38.
Lotterhos KE. The effect of neutral recombination variation on genome scans for selection. G3-Genes Genom Genet. 2019;9(6):1851–67.
Lovell JT, MacQueen AH, Mamidi S, Bonnette J, Jenkins J, Napier JD, et al. Genomic mechanisms of climate adaptation in polyploid bioenergy switchgrass. Nature. 2021;590(7846):438–44.
Todesco M, Owens GL, Bercovich N, Légaré J-S, Soudi S, Burge DO, et al. Massive haplotypes underlie ecotypic differentiation in sunflowers. Nature. 2020;584(7822):602–7.
Revolinski SR, Maughan PJ, Coleman CE, Burke IC. Preadapted to adapt: Underpinnings of adaptive plasticity revealed by the downy brome genome. Commun Biol. 2023;6(1):326.
Kuester A, Conner JK, Culley T, Baucom RS. How weeds emerge: a taxonomic and trait-based examination using United States data. New Phytol. 2014;202(3):1055–68.
Arnaud JF, Fénart S, Cordellier M, Cuguen J. Populations of weedy crop-wild hybrid beets show contrasting variation in mating system and population genetic structure. Evol Appl. 2010;3(3):305–18.
Ellstrand NC, Schierenbeck KA. Hybridization as a stimulus for the evolution of invasiveness in plants? Proc Natl Acad Sci U S A. 2000;97(13):7043–50.
Nakabayashi K, Leubner-Metzger G. Seed dormancy and weed emergence: from simulating environmental change to understanding trait plasticity, adaptive evolution, and population fitness. J Exp Bot. 2021;72(12):4181–5.
Busi R, Yu Q, Barrett-Lennard R, Powles S. Long distance pollen-mediated flow of herbicide resistance genes in Lolium rigidum . Theor Appl Genet. 2008;117(8):1281–90.
Délye C, Clément JAJ, Pernin F, Chauvel B, Le Corre V. High gene flow promotes the genetic homogeneity of arable weed populations at the landscape level. Basic Appl Ecol. 2010;11(6):504–12.
Roumet M, Noilhan C, Latreille M, David J, Muller MH. How to escape from crop-to-weed gene flow: phenological variation and isolation-by-time within weedy sunflower populations. New Phytol. 2013;197(2):642–54.
Moghadam SH, Alebrahim MT, Mohebodini M, MacGregor DR. Genetic variation of Amaranthus retroflexus L. and Chenopodium album L. (Amaranthaceae) suggests multiple independent introductions into Iran. Front Plant Sci. 2023;13:1024555.
Muller M-H, Latreille M, Tollon C. The origin and evolution of a recent agricultural weed: population genetic diversity of weedy populations of sunflower ( Helianthus annuus L.) in Spain and France. Evol Appl. 2011;4(3):499–514.
Wesse C, Welk E, Hurka H, Neuffer B. Geographical pattern of genetic diversity in Capsella bursa-pastoris (Brassicaceae) -A global perspective. Ecol Evol. 2021;11(1):199–213.
Fraimout A, Debat V, Fellous S, Hufbauer RA, Foucaud J, Pudlo P, et al. Deciphering the routes of invasion of Drosophila suzukii by means of ABC random forest. Mol Biol Evol. 2017;34(4):980–96.
CAS PubMed PubMed Central Google Scholar
Battlay P, Wilson J, Bieker VC, Lee C, Prapas D, Petersen B, et al. Large haploblocks underlie rapid adaptation in the invasive weed Ambrosia artemisiifolia . Nat Commun. 2023;14(1):1717.
van Boheemen LA, Hodgins KA. Rapid repeatable phenotypic and genomic adaptation following multiple introductions. Mol Ecol. 2020;29(21):4102–17.
Putra A, Hodgins K, Fournier-Level A. Assessing the invasive potential of different source populations of ragweed ( Ambrosia artemisiifolia L.) through genomically-informed species distribution modelling. Authorea. 2023;17(1):e13632.
Google Scholar
Bourguet D, Delmotte F, Franck P, Guillemaud T, Reboud X, Vacher C, et al. Heterogeneity of selection and the evolution of resistance. Trends Ecol Evol. 2013;28(2):110–8.
The International Herbicide-Resistant Weed Database. www.weedscience.org . Accessed 20 June 2023.
Powles S. Herbicide discovery through innovation and diversity. Adv Weed Sci. 2022;40(spe1):e020220074.
Murphy BP, Tranel PJ. Target-site mutations conferring herbicide resistance. Plants (Basel). 2019;8(10):382.
Gaines TA, Duke SO, Morran S, Rigon CAG, Tranel PJ, Küpper A, et al. Mechanisms of evolved herbicide resistance. J Biol Chem. 2020;295(30):10307–30.
Lonhienne T, Cheng Y, Garcia MD, Hu SH, Low YS, Schenk G, et al. Structural basis of resistance to herbicides that target acetohydroxyacid synthase. Nat Commun. 2022;13(1):3368.
Comont D, MacGregor DR, Crook L, Hull R, Nguyen L, Freckleton RP, et al. Dissecting weed adaptation: fitness and trait correlations in herbicide-resistant Alopecurus myosuroides . Pest Manag Sci. 2022;78(7):3039–50.
Neve P. Simulation modelling to understand the evolution and management of glyphosate resistance in weeds. Pest Manag Sci. 2008;64(4):392–401.
Torra J, Alcántara-de la Cruz R. Molecular mechanisms of herbicide resistance in weeds. Genes (Basel). 2022;13(11):2025.
Délye C, Gardin JAC, Boucansaud K, Chauvel B, Petit C. Non-target-site-based resistance should be the centre of attention for herbicide resistance research: Alopecurus myosuroides as an illustration. Weed Res. 2011;51(5):433–7.
Chandra S, Leon RG. Genome-wide evolutionary analysis of putative non-specific herbicide resistance genes and compilation of core promoters between monocots and dicots. Genes (Basel). 2022;13(7):1171.
Margaritopoulou T, Tani E, Chachalis D, Travlos I. Involvement of epigenetic mechanisms in herbicide resistance: the case of Conyza canadensis . Agriculture. 2018;8(1):17.
Pan L, Guo Q, Wang J, Shi L, Yang X, Zhou Y, et al. CYP81A68 confers metabolic resistance to ALS and ACCase-inhibiting herbicides and its epigenetic regulation in Echinochloa crus-galli . J Hazard Mater. 2022;428:128225.
Sen MK, Hamouzová K, Košnarová P, Roy A, Soukup J. Herbicide resistance in grass weeds: Epigenetic regulation matters too. Front Plant Sci. 2022;13:1040958.
Han H, Yu Q, Beffa R, González S, Maiwald F, Wang J, et al. Cytochrome P450 CYP81A10v7 in Lolium rigidum confers metabolic resistance to herbicides across at least five modes of action. Plant J. 2021;105(1):79–92.
Kubis GC, Marques RZ, Kitamura RS, Barroso AA, Juneau P, Gomes MP. Antioxidant enzyme and Cytochrome P450 activities are involved in horseweed ( Conyza sumatrensis ) resistance to glyphosate. Stress. 2023;3(1):47–57.
Qiao Y, Zhang N, Liu J, Yang H. Interpretation of ametryn biodegradation in rice based on joint analyses of transcriptome, metabolome and chemo-characterization. J Hazard Mater. 2023;445:130526.
Rouse CE, Roma-Burgos N, Barbosa Martins BA. Physiological assessment of non–target site restistance in multiple-resistant junglerice ( Echinochloa colona ). Weed Sci. 2019;67(6):622–32.
Abou-Khater L, Maalouf F, Jighly A, Alsamman AM, Rubiales D, Rispail N, et al. Genomic regions associated with herbicide tolerance in a worldwide faba bean ( Vicia faba L.) collection. Sci Rep. 2022;12(1):158.
Gupta S, Harkess A, Soble A, Van Etten M, Leebens-Mack J, Baucom RS. Interchromosomal linkage disequilibrium and linked fitness cost loci associated with selection for herbicide resistance. New Phytol. 2023;238(3):1263–77.
Kreiner JM, Tranel PJ, Weigel D, Stinchcombe JR, Wright SI. The genetic architecture and population genomic signatures of glyphosate resistance in Amaranthus tuberculatus . Mol Ecol. 2021;30(21):5373–89.
Parcharidou E, Dücker R, Zöllner P, Ries S, Orru R, Beffa R. Recombinant glutathione transferases from flufenacet-resistant black-grass ( Alopecurus myosuroides Huds.) form different flufenacet metabolites and differ in their interaction with pre- and post-emergence herbicides. Pest Manag Sci. 2023;79(9):3376–86.
Békés M, Langley DR, Crews CM. PROTAC targeted protein degraders: the past is prologue. Nat Rev Drug Discov. 2022;21(3):181–200.
Acuner Ozbabacan SE, Engin HB, Gursoy A, Keskin O. Transient protein-protein interactions. Protein Eng Des Sel. 2011;24(9):635–48.
Lu H, Zhou Q, He J, Jiang Z, Peng C, Tong R, et al. Recent advances in the development of protein–protein interactions modulators: mechanisms and clinical trials. Signal Transduct Target Ther. 2020;5(1):213.
Benson CW, Sheltra MR, Maughan PJ, Jellen EN, Robbins MD, Bushman BS, et al. Homoeologous evolution of the allotetraploid genome of Poa annua L. BMC Genom. 2023;24(1):350.
Robbins MD, Bushman BS, Huff DR, Benson CW, Warnke SE, Maughan CA, et al. Chromosome-scale genome assembly and annotation of allotetraploid annual bluegrass ( Poa annua L.). Genome Biol Evol. 2022;15(1):evac180.
Montgomery JS, Giacomini D, Waithaka B, Lanz C, Murphy BP, Campe R, et al. Draft genomes of Amaranthus tuberculatus , Amaranthus hybridus and Amaranthus palmeri . Genome Biol Evol. 2020;12(11):1988–93.
Jeschke MR, Tranel PJ, Rayburn AL. DNA content analysis of smooth pigweed ( Amaranthus hybridus ) and tall waterhemp ( A. tuberculatus ): implications for hybrid detection. Weed Sci. 2003;51(1):1–3.
Rayburn AL, McCloskey R, Tatum TC, Bollero GA, Jeschke MR, Tranel PJ. Genome size analysis of weedy Amaranthus species. Crop Sci. 2005;45(6):2557–62.
Laforest M, Martin SL, Bisaillon K, Soufiane B, Meloche S, Tardif FJ, et al. The ancestral karyotype of the Heliantheae Alliance, herbicide resistance, and human allergens: Insights from the genomes of common and giant ragweed. Plant Genome . 2024;e20442. https://doi.org/10.1002/tpg2.20442 .
Mulligan GA. Chromosome numbers of Canadian weeds. I Canad J Bot. 1957;35(5):779–89.
Meyer L, Causse R, Pernin F, Scalone R, Bailly G, Chauvel B, et al. New gSSR and EST-SSR markers reveal high genetic diversity in the invasive plant Ambrosia artemisiifolia L. and can be transferred to other invasive Ambrosia species. PLoS One. 2017;12(5):e0176197.
Pustahija F, Brown SC, Bogunić F, Bašić N, Muratović E, Ollier S, et al. Small genomes dominate in plants growing on serpentine soils in West Balkans, an exhaustive study of 8 habitats covering 308 taxa. Plant Soil. 2013;373(1):427–53.
Kubešová M, Moravcova L, Suda J, Jarošík V, Pyšek P. Naturalized plants have smaller genomes than their non-invading relatives: a flow cytometric analysis of the Czech alien flora. Preslia. 2010;82(1):81–96.
Thébaud C, Abbott RJ. Characterization of invasive Conyza species (Asteraceae) in Europe: quantitative trait and isozyme analysis. Am J Bot. 1995;82(3):360–8.
Garcia S, Hidalgo O, Jakovljević I, Siljak-Yakovlev S, Vigo J, Garnatje T, et al. New data on genome size in 128 Asteraceae species and subspecies, with first assessments for 40 genera, 3 tribes and 2 subfamilies. Plant Biosyst. 2013;147(4):1219–27.
Zhao X, Yi L, Ren Y, Li J, Ren W, Hou Z, et al. Chromosome-scale genome assembly of the yellow nutsedge ( Cyperus esculentus ). Genome Biol Evol. 2023;15(3):evad027.
Bennett MD, Leitch IJ, Hanson L. DNA amounts in two samples of angiosperm weeds. Ann Bot. 1998;82:121–34.
Schulz-Schaeffer J, Gerhardt S. Cytotaxonomic analysis of the Euphorbia spp. (leafy spurge) complex. II: Comparative study of the chromosome morphology. Biol Zentralbl. 1989;108(1):69–76.
Schaeffer JR, Gerhardt S. The impact of introgressive hybridization on the weediness of leafy spurge. Leafy Spurge Symposium. 1989;1989:97–105.
Bai C, Alverson WS, Follansbee A, Waller DM. New reports of nuclear DNA content for 407 vascular plant taxa from the United States. Ann Bot. 2012;110(8):1623–9.
Aarestrup JR, Karam D, Fernandes GW. Chromosome number and cytogenetics of Euphorbia heterophylla L. Genet Mol Res. 2008;7(1):217–22.
Wang L, Sun X, Peng Y, Chen K, Wu S, Guo Y, et al. Genomic insights into the origin, adaptive evolution, and herbicide resistance of Leptochloa chinensis , a devastating tetraploid weedy grass in rice fields. Mol Plant. 2022;15(6):1045–58.
Paril J, Pandey G, Barnett EM, Rane RV, Court L, Walsh T, et al. Rounding up the annual ryegrass genome: high-quality reference genome of Lolium rigidum . Front Genet. 2022;13:1012694.
Weiss-Schneeweiss H, Greilhuber J, Schneeweiss GM. Genome size evolution in holoparasitic Orobanche (Orobanchaceae) and related genera. Am J Bot. 2006;93(1):148–56.
Towers G, Mitchell J, Rodriguez E, Bennett F, Subba Rao P. Biology & chemistry of Parthenium hysterophorus L., a problem weed in India. Biol Rev. 1977;48:65–74.
CAS Google Scholar
Moghe GD, Hufnagel DE, Tang H, Xiao Y, Dworkin I, Town CD, et al. Consequences of whole-genome triplication as revealed by comparative genomic analyses of the wild radish ( Raphanus raphanistrum ) and three other Brassicaceae species. Plant Cell. 2014;26(5):1925–37.
Zhang X, Liu T, Wang J, Wang P, Qiu Y, Zhao W, et al. Pan-genome of Raphanus highlights genetic variation and introgression among domesticated, wild, and weedy radishes. Mol Plant. 2021;14(12):2032–55.
Chytrý M, Danihelka J, Kaplan Z, Wild J, Holubová D, Novotný P, et al. Pladias database of the Czech flora and vegetation. Preslia. 2021;93(1):1–87.
Patterson EL, Pettinga DJ, Ravet K, Neve P, Gaines TA. Glyphosate resistance and EPSPS gene duplication: Convergent evolution in multiple plant species. J Hered. 2018;109(2):117–25.
Jugulam M, Niehues K, Godar AS, Koo DH, Danilova T, Friebe B, et al. Tandem amplification of a chromosomal segment harboring 5-enolpyruvylshikimate-3-phosphate synthase locus confers glyphosate resistance in Kochia scoparia . Plant Physiol. 2014;166(3):1200–7.
Patterson EL, Saski CA, Sloan DB, Tranel PJ, Westra P, Gaines TA. The draft genome of Kochia scoparia and the mechanism of glyphosate resistance via transposon-mediated EPSPS tandem gene duplication. Genome Biol Evol. 2019;11(10):2927–40.
Zhang C, Johnson N, Hall N, Tian X, Yu Q, Patterson E. Subtelomeric 5-enolpyruvylshikimate-3-phosphate synthase ( EPSPS ) copy number variation confers glyphosate resistance in Eleusine indica . Nat Commun. 2023;14:4865.
Koo D-H, Molin WT, Saski CA, Jiang J, Putta K, Jugulam M, et al. Extrachromosomal circular DNA-based amplification and transmission of herbicide resistance in crop weed Amaranthus palmeri . Proc Natl Acad Sci U S A. 2018;115(13):3332–7.
Molin WT, Yaguchi A, Blenner M, Saski CA. The eccDNA Replicon: A heritable, extranuclear vehicle that enables gene amplification and glyphosate resistance in Amaranthus palmeri . Plant Cell. 2020;32(7):2132–40.
Jugulam M. Can non-Mendelian inheritance of extrachromosomal circular DNA-mediated EPSPS gene amplification provide an opportunity to reverse resistance to glyphosate? Weed Res. 2021;61(2):100–5.
Kreiner JM, Giacomini DA, Bemm F, Waithaka B, Regalado J, Lanz C, et al. Multiple modes of convergent adaptation in the spread of glyphosate-resistant Amaranthus tuberculatus . Proc Natl Acad Sci U S A. 2019;116(42):21076–84.
Cai L, Comont D, MacGregor D, Lowe C, Beffa R, Neve P, et al. The blackgrass genome reveals patterns of non-parallel evolution of polygenic herbicide resistance. New Phytol. 2023;237(5):1891–907.
Chen K, Yang H, Peng Y, Liu D, Zhang J, Zhao Z, et al. Genomic analyses provide insights into the polyploidization-driven herbicide adaptation in Leptochloa weeds. Plant Biotechnol J. 2023;21(8):1642–58.
Ohadi S, Hodnett G, Rooney W, Bagavathiannan M. Gene flow and its consequences in Sorghum spp. Crit Rev Plant Sci. 2017;36(5–6):367–85.
Renzi JP, Coyne CJ, Berger J, von Wettberg E, Nelson M, Ureta S, et al. How could the use of crop wild relatives in breeding increase the adaptation of crops to marginal environments? Front Plant Sci. 2022;13:886162.
Ward SM, Cousens RD, Bagavathiannan MV, Barney JN, Beckie HJ, Busi R, et al. Agricultural weed research: a critique and two proposals. Weed Sci. 2014;62(4):672–8.
Evans JA, Tranel PJ, Hager AG, Schutte B, Wu C, Chatham LA, et al. Managing the evolution of herbicide resistance. Pest Manag Sci. 2016;72(1):74–80.
International Weed Genomics Consortium Website. https://www.weedgenomics.org . Accessed 20 June 2023.
WeedPedia Database. https://weedpedia.weedgenomics.org/ . Accessed 20 June 2023.
Hall N, Chen J, Matzrafi M, Saski CA, Westra P, Gaines TA, et al. FHY3/FAR1 transposable elements generate adaptive genetic variation in the Bassia scoparia genome. bioRxiv . 2023; DOI: https://doi.org/10.1101/2023.05.26.542497 .
Jarvis DE, Sproul JS, Navarro-Domínguez B, Krak K, Jaggi K, Huang Y-F, et al. Chromosome-scale genome assembly of the hexaploid Taiwanese goosefoot “Djulis” ( Chenopodium formosanum ). Genome Biol Evol. 2022;14(8):evac120.
Ferreira LAI, de Oliveira RS, Jr., Constantin J, Brunharo C. Evolution of ACCase-inhibitor resistance in Chloris virgata is conferred by a Trp2027Cys mutation in the herbicide target site. Pest Manag Sci. 2023;79(12):5220–9.
Laforest M, Martin SL, Bisaillon K, Soufiane B, Meloche S, Page E. A chromosome-scale draft sequence of the Canada fleabane genome. Pest Manag Sci. 2020;76(6):2158–69.
Guo L, Qiu J, Ye C, Jin G, Mao L, Zhang H, et al. Echinochloa crus-galli genome analysis provides insight into its adaptation and invasiveness as a weed. Nat Commun. 2017;8(1):1031.
Sato MP, Iwakami S, Fukunishi K, Sugiura K, Yasuda K, Isobe S, et al. Telomere-to-telomere genome assembly of an allotetraploid pernicious weed, Echinochloa phyllopogon . DNA Res. 2023;30(5):dsad023.
Stein JC, Yu Y, Copetti D, Zwickl DJ, Zhang L, Zhang C, et al. Genomes of 13 domesticated and wild rice relatives highlight genetic conservation, turnover and innovation across the genus Oryza . Nat Genet. 2018;50(2):285–96.
Wu D, Xie L, Sun Y, Huang Y, Jia L, Dong C, et al. A syntelog-based pan-genome provides insights into rice domestication and de-domestication. Genome Biol. 2023;24(1):179.
Wang Z, Huang S, Yang Z, Lai J, Gao X, Shi J. A high-quality, phased genome assembly of broomcorn millet reveals the features of its subgenome evolution and 3D chromatin organization. Plant Commun. 2023;4(3):100557.
Mao Q, Huff DR. The evolutionary origin of Poa annua L. Crop Sci. 2012;52(4):1910–22.
Benson CW, Sheltra MR, Maughan JP, Jellen EN, Robbins MD, Bushman BS, et al. Homoeologous evolution of the allotetraploid genome of Poa annua L. Res Sq. 2023. https://doi.org/10.21203/rs.3.rs-2729084/v1 .
Brunharo C, Benson CW, Huff DR, Lasky JR. Chromosome-scale genome assembly of Poa trivialis and population genomics reveal widespread gene flow in a cool-season grass seed production system. Plant Direct. 2024;8(3):e575.
Mo C, Wu Z, Shang X, Shi P, Wei M, Wang H, et al. Chromosome-level and graphic genomes provide insights into metabolism of bioactive metabolites and cold-adaption of Pueraria lobata var. montana . DNA Research. 2022;29(5):dsac030.
Thielen PM, Pendleton AL, Player RA, Bowden KV, Lawton TJ, Wisecaver JH. Reference genome for the highly transformable Setaria viridis ME034V. G3 (Bethesda, Md). 2020;10(10):3467–78.
Yoshida S, Kim S, Wafula EK, Tanskanen J, Kim Y-M, Honaas L, et al. Genome sequence of Striga asiatica provides insight into the evolution of plant parasitism. Curr Biol. 2019;29(18):3041–52.
Qiu S, Bradley JM, Zhang P, Chaudhuri R, Blaxter M, Butlin RK, et al. Genome-enabled discovery of candidate virulence loci in Striga hermonthica , a devastating parasite of African cereal crops. New Phytol. 2022;236(2):622–38.
Nunn A, Rodríguez-Arévalo I, Tandukar Z, Frels K, Contreras-Garrido A, Carbonell-Bejerano P, et al. Chromosome-level Thlaspi arvense genome provides new tools for translational research and for a newly domesticated cash cover crop of the cooler climates. Plant Biotechnol J. 2022;20(5):944–63.
USDA-ARS Germplasm Resources Information Network (GRIN). https://www.ars-grin.gov/ . Accessed 20 June 2023.
Buck M, Hamilton C. The Nagoya Protocol on access to genetic resources and the fair and equitable sharing of benefits arising from their utilization to the convention on biological diversity. RECIEL. 2011;20(1):47–61.
Chauhan BS, Matloob A, Mahajan G, Aslam F, Florentine SK, Jha P. Emerging challenges and opportunities for education and research in weed science. Front Plant Sci. 2017;8:1537.
Shah S, Lonhienne T, Murray CE, Chen Y, Dougan KE, Low YS, et al. Genome-guided analysis of seven weed species reveals conserved sequence and structural features of key gene targets for herbicide development. Front Plant Sci. 2022;13:909073.
International Weed Genomics Consortium Training Resources. https://www.weedgenomics.org/training-resources/ . Accessed 20 June 2023.
Blackford S. Harnessing the power of communities: career networking strategies for bioscience PhD students and postdoctoral researchers. FEMS Microbiol Lett. 2018;365(8):fny033.
Pender M, Marcotte DE, Sto Domingo MR, Maton KI. The STEM pipeline: The role of summer research experience in minority students’ Ph.D. aspirations. Educ Policy Anal Arch. 2010;18(30):1–36.
PubMed PubMed Central Google Scholar
Burke A, Okrent A, Hale K. The state of U.S. science and engineering 2022. Foundation NS. https://ncses.nsf.gov/pubs/nsb20221 . 2022.
Wu J-Y, Liao C-H, Cheng T, Nian M-W. Using data analytics to investigate attendees’ behaviors and psychological states in a virtual academic conference. Educ Technol Soc. 2021;24(1):75–91.
Download references
Peer review information
Wenjing She was the primary editor of this article and managed its editorial process and peer review in collaboration with the rest of the editorial team.
The International Weed Genomics Consortium is supported by BASF SE, Bayer AG, Syngenta Ltd, Corteva Agriscience, CropLife International (Global Herbicide Resistance Action Committee), the Foundation for Food and Agriculture Research (Award DSnew-0000000024), and two conference grants from USDA-NIFA (Award numbers 2021–67013-33570 and 2023-67013-38785).
Author information
Authors and affiliations.
Department of Agricultural Biology, Colorado State University, 1177 Campus Delivery, Fort Collins, CO, 80523, USA
Jacob Montgomery, Sarah Morran & Todd A. Gaines
Protecting Crops and the Environment, Rothamsted Research, Harpenden, Hertfordshire, UK
Dana R. MacGregor
Department of Crop, Soil, and Environmental Sciences, Auburn University, Auburn, AL, USA
J. Scott McElroy
Department of Plant and Environmental Sciences, University of Copenhagen, Taastrup, Denmark
Paul Neve & Célia Neto
IFEVA-Conicet-Department of Ecology, University of Buenos Aires, Buenos Aires, Argentina
Martin M. Vila-Aiub & Maria Victoria Sandoval
Department of Ecology, Faculty of Agronomy, University of Buenos Aires, Buenos Aires, Argentina
Analia I. Menéndez
Department of Botany, The University of British Columbia, Vancouver, BC, Canada
Julia M. Kreiner
Institute of Crop Sciences, Zhejiang University, Hangzhou, China
Longjiang Fan
Department of Biology, University of Massachusetts Amherst, Amherst, MA, USA
Ana L. Caicedo
Department of Plant and Wildlife Sciences, Brigham Young University, Provo, UT, USA
Peter J. Maughan
Bayer AG, Weed Control Research, Frankfurt, Germany
Bianca Assis Barbosa Martins, Jagoda Mika, Alberto Collavo & Bodo Peters
Department of Crop Sciences, Federal University of Rio Grande Do Sul, Porto Alegre, Rio Grande Do Sul, Brazil
Aldo Merotto Jr.
Department of Soil and Crop Sciences, Texas A&M University, College Station, TX, USA
Nithya K. Subramanian & Muthukumar V. Bagavathiannan
Department of Plant, Soil and Microbial Sciences, Michigan State University, East Lansing, MI, USA
Luan Cutti & Eric L. Patterson
Department of Agronomy, Kansas State University, Manhattan, KS, USA
Md. Mazharul Islam & Mithila Jugulam
Department of Plant Pathology, Kansas State University, Manhattan, KS, USA
Bikram S. Gill
Crop Protection Discovery and Development, Corteva Agriscience, Indianapolis, IN, USA
Robert Cicchillo, Roger Gast & Neeta Soni
Genome Center of Excellence, Corteva Agriscience, Johnston, IA, USA
Terry R. Wright, Gina Zastrow-Hayes, Gregory May, Kevin Fengler & Victor Llaca
School of Agriculture, Food and Wine, University of Adelaide, Glen Osmond, South Australia, Australia
Jenna M. Malone
Jealott’s Hill International Research Centre, Syngenta Ltd, Bracknell, Berkshire, UK
Deepmala Sehgal, Shiv Shankhar Kaundun & Richard P. Dale
Department of Plant and Soil Sciences, University of Pretoria, Pretoria, South Africa
Barend Juan Vorster
BASF SE, Ludwigshafen Am Rhein, Germany
Jens Lerchl
Department of Crop Sciences, University of Illinois, Urbana, IL, USA
Patrick J. Tranel
Senior Scientist Consultant, Herbicide Resistance Action Committee / CropLife International, Liederbach, Germany
Roland Beffa
School of BioSciences, University of Melbourne, Parkville, VIC, Australia
Alexandre Fournier-Level
You can also search for this author in PubMed Google Scholar
Contributions
JMo and TG conceived and outlined the article. TG, DM, EP, RB, JSM, PJT, MJ wrote grants to obtain funding. MMI, BSG, and MJ performed mitotic chromosome visualization. VL performed sequencing. VL and KF assembled the genomes. LC and ELP annotated the genomes. JMo, SM, DRM, JSM, PN, CN, MV, MVS, AIM, JMK, LF, ALC, PJM, BABM, JMi, AC, MVB, LC, AFL, and ELP wrote the first draft of the article. All authors edited the article and improved the final version.
Corresponding author
Correspondence to Todd A. Gaines .
Ethics declarations
Ethics approval and consent to participate.
Ethical approval is not applicable for this article.
Competing interests
Some authors work for commercial agricultural companies (BASF, Bayer, Corteva Agriscience, or Syngenta) that develop and sell weed control products.
Additional information
Publisher’s note.
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
13059_2024_3274_moesm1_esm.docx.
Additional file 1. List of completed and in-progress genome assemblies of weed species pollinated by insects (Table S1).
13059_2024_3274_MOESM2_ESM.docx
Additional file 2. Methods and results for visualizing and counting the metaphase chromosomes of hexaploid Avena fatua (Fig S1); diploid Lolium rigidum (Fig S2); tetraploid Phalaris minor (Fig S3); and tetraploid Salsola tragus (Fig S4).
Additional file 3. Review history.
Rights and permissions.
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/ . The Creative Commons Public Domain Dedication waiver ( http://creativecommons.org/publicdomain/zero/1.0/ ) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
Reprints and permissions
About this article
Cite this article.
Montgomery, J., Morran, S., MacGregor, D.R. et al. Current status of community resources and priorities for weed genomics research. Genome Biol 25 , 139 (2024). https://doi.org/10.1186/s13059-024-03274-y
Download citation
Received : 11 July 2023
Accepted : 13 May 2024
Published : 27 May 2024
DOI : https://doi.org/10.1186/s13059-024-03274-y
Share this article
Anyone you share the following link with will be able to read this content:
Sorry, a shareable link is not currently available for this article.
Provided by the Springer Nature SharedIt content-sharing initiative
- Weed science
- Reference genomes
- Rapid adaptation
- Herbicide resistance
- Public resources
Genome Biology
ISSN: 1474-760X
- Submission enquiries: [email protected]
- General enquiries: [email protected]
IMAGES
VIDEO
COMMENTS
Learn about the latest stress statistics, sources, and management techniques in the US and worldwide. Explore the impact of stress on health, relationships, work, and coping mechanisms.
LIFE STRESS, ANXIETY, AND DEPRESSION . It is well known that first depressive episodes often develop following the occurrence of a major negative life event (Paykel 2001).Furthermore, there is evidence that stressful life events are causal for the onset of depression (see Hammen 2005, Kendler et al. 1999).A study of 13,006 patients in Denmark, with first psychiatric admissions diagnosed with ...
Stress and the Brain Function Complications. For a long time, researchers suggested that hormones have receptors just in the peripheral tissues and do not gain access to the central nervous system (CNS) (Lupien and Lepage, 2001[]).However, observations have demonstrated the effect of anti-inflammatory drugs (which are considered synthetic hormones) on behavioral and cognitive disorders and the ...
Original research, reviews, and perspectives are presented on how stress affects development, metabolism, and various cellular and organ level processes of physiology. We hope this issue contributes to an emerging field and highlights the importance of an interdisciplinary approach to understanding the wide implications of stress.
Learn about stress, its effects on the body and mind, and how to cope with it. Find resources on stress research, statistics, tips, and books from APA.
The cumulative science linking stress to negative health outcomes is vast. Stress can affect health directly, through autonomic and neuroendocrine responses, but also indirectly, through changes in health behaviors. In this review, we present a brief overview of (a) why we should be interested in stress in the context of health; (b) the stress response and allostatic load; (c) some of the key ...
The legacy of Hans Selye and the origins of stress research: a retrospective 75 years after his landmark brief "letter" to the editor of Nature. Stress 15 , 472-478 (2012). CAS PubMed Google ...
Stress & Health is an international forum for disseminating cutting-edge theoretical and empirical research that significantly advances understanding of the relationship between stress and health and well-being in humans. Fast online publication with your accepted manuscript publishing online within a week, with manuscript transfer option.
Coinciding with WASAD's 3rd International Congress held in September 2021 in Vienna, Austria, this journal publishes a Special Issue encompassing state-of-the art research in the field of stress and anxiety. This special issue collects answers to a number of important questions that need to be addressed in current and future research.
Learn how stress affects the body, mind, and society, and what coping strategies can help you manage it. Find out how stress can be damaging, protective, or transformative depending on the situation and support.
While Selye focused mainly on the stressor and the (patho) physiology of the stress response, the research of Levine and others emphasized that stress is about the processing of the individual experience of the stressor and the ability to cope. Thus, the most severe stress is a psychological condition characterized by lack of information to ...
One of the leading frameworks in work stress research that is used to guide organizational interventions is the person and environment (P-E) fit (French & Caplan, 1972). Its precursor is the University of Michigan Institute for Social Research's (ISR) role stress model (Kahn, Wolfe, Quinn, Snoek, & Rosenthal, 1964) and Lewin's Field Theory.
80% of US workers experience work stress because of ineffective company communications. 39% of North American employees report their workload the main source of the work stress. 49% of 18 - 24 year olds who report high levels of stress felt comparing themselves to others is a stressor. 71% of US adults with private health insurance say the ...
The AIS Seal and credentials inform the public that the certificate holder commands advanced knowledge of the latest stress research and stress management techniques. For physicians and other healthcare practitioners it designates your practice as an advanced treatment center for stress related illnesses.
In sum, researchers should identify the type or types of stress that are most relevant to their research question and sample. Stress measure selection should then be refined based on characteristics of the stressor and/or stress response that the researcher intends to measure, such as the timescale, the type(s) of stress responses the ...
Stress is a multidimensional construct that comprises exposure to events, perceptions of stress, and physiological responses to stress. Research consistently demonstrates a strong association between stress and a myriad of physical and mental health concerns, resulting in a pervasive and interdisciplinary agreement on the importance of investigating the relationship between stress and health.
Methodology. The 2020 Stress in America™ survey was conducted online within the United States by The Harris Poll on behalf of the American Psychological Association between Aug. 4 and Aug. 26, 2020, among 3,409 adults age 18+ who reside in the U.S. Interviews were conducted in English and Spanish.
Learn how stress affects health and how complementary and integrative health interventions can help manage it. See examples of NCCIH-funded studies on stress management for people with chronic diseases and the benefits of yoga for diabetes.
Research has long shown that exercise reduces heart disease risk, but a new study finds the connection between the two may be a decrease in the brain's stress signals. CNN values your feedback 1.
Constantly announcing to co-workers how swamped you are at the office isn't likely to impress your colleagues — in fact, it may have the opposite effect, new research shows. "Stress bragging" or ...
Reducing your stress levels can not only make you feel better right now, but may also protect your health long-term. Several research studies have demonstrated, for example, that interventions to improve psychological health can have a beneficial impact on cardiovascular health. As a result, researchers recommend boosting your positive affect ...
Research shows how stress activates neurons that disrupt sleep. Dec 13, 2023. Why do embarrassing memories seem to surface at night? Jun 27, 2022. Brain wave activity associated with circadian ...
2. The need for stress research during the current COVID-19 pandemic. In recent years, the prevalence of stress-related mental disorders has been following an upward trend (Baxter et al., 2014; Cohen and Janicki-Deverts, 2012; DeVries and Wilkerson, 2003), causing both individual burden and financial and social problems for society as a whole (Hassard et al., 2018; Trautmann et al., 2016).
This process allows cells to respond rapidly to stress by altering RNA molecules within a cell. As such, this breakthrough discovery advances the understanding of how malaria parasites respond to drug-induced stress and develop resistance, and paves the way for the development of new drugs to combat resistance. ... The research sets the ...
Squamous promoter binding protein-like (SPL) genes encode plant-specific transcription factors (TFs) that play essential roles in modulating plant growth, development, and stress response. Pea (Pisum sativum L.) is a coarse grain crop of great importance in food production, biodiversity conservation and molecular genetic research, providing genetic information and nutritional resources for ...
Research on work stress and physical health. Epidemiological studies consistently demonstrate associations between high work stress and worse physical and mental health. One of the most widely studied models of work stress is job strain, which is a combination of high demands ...
Studies have suggested that endoplasmic reticulum stress (ERS) is involved in neurological dysfunction and that electroacupuncture (EA) attenuates neuropathic pain (NP) via undefined pathways. However, the role of ERS in the anterior cingulate cortex (ACC) in NP and the effect of EA on ERS in the ACC have not yet been investigated. In this study, an NP model was established by chronic ...
Weeds are attractive models for basic and applied research due to their impacts on agricultural systems and capacity to swiftly adapt in response to anthropogenic selection pressures. Currently, a lack of genomic information precludes research to elucidate the genetic basis of rapid adaptation for important traits like herbicide resistance and stress tolerance and the effect of evolutionary ...