Thank you for visiting nature.com. You are using a browser version with limited support for CSS. To obtain the best experience, we recommend you use a more up to date browser (or turn off compatibility mode in Internet Explorer). In the meantime, to ensure continued support, we are displaying the site without styles and JavaScript.
- View all journals
- Explore content
- About the journal
- Publish with us
- Sign up for alerts
- Published: 01 August 2005

Forty-five years of split-brain research and still going strong
- Michael S. Gazzaniga 1
Nature Reviews Neuroscience volume 6 , pages 653–659 ( 2005 ) Cite this article
10k Accesses
276 Citations
32 Altmetric
Metrics details
Forty-five years ago, Roger Sperry, Joseph Bogen and I embarked on what are now known as the modern split-brain studies. These experiments opened up new frontiers in brain research and gave rise to much of what we know about hemispheric specialization and integration. The latest developments in split-brain research build on the groundwork laid by those early studies. Split-brain methodology, on its own and in conjunction with neuroimaging, has yielded insights into the remarkable regional specificity of the corpus callosum as well as into the integrative role of the callosum in the perception of causality and in our perception of an integrated sense of self.
This is a preview of subscription content, access via your institution
Relevant articles
Open Access articles citing this article.
A review of combined functional neuroimaging and motion capture for motor rehabilitation
- Emanuel A. Lorenz
- , Xiaomeng Su
- & Nina Skjæret-Maroni
Journal of NeuroEngineering and Rehabilitation Open Access 03 January 2024
A frontal transcallosal inhibition loop mediates interhemispheric balance in visuospatial processing
- Yanjie Wang
- , Zhaonan Chen
- … Siyu Zhang
Nature Communications Open Access 25 August 2023
Mapping lesion, structural disconnection, and functional disconnection to symptoms in semantic aphasia
- Nicholas E. Souter
- , Xiuyi Wang
- … Elizabeth Jefferies
Brain Structure and Function Open Access 04 July 2022
Access options
Subscribe to this journal
Receive 12 print issues and online access
176,64 € per year
only 14,72 € per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout
Lashley, K. S. In search of the engram. Symp. Soc. Exp. Biol. 4 , 454–482 (1950).
Google Scholar
Hebb, D. O. The Organization of Behavior: a Neuropsychological Theory (Wiley, New York, USA, 1949).
Sperry, R. W. Chemoaffinity in the orderly growth of nerve fiber patterns and connections. Proc. Natl Acad. Sci. USA 50 , 703–710 (1963).
CAS PubMed Google Scholar
Weiss, P. A. In vitro experiments on the factors determining the course of the outgrowing nerve fiber. J. Exp. Zool. 68 , 393–448 (1934).
Van Wagenen, W. P & Herren, R. Y. Surgical division of commissural pathways in the corpus callosum: relation to spread of an epileptic attack. Arch. Neurol. Psychiatry 44 , 740–759 (1940).
Bogen, J. E. & Vogel, P. J. Cerebral commissurotomy in man. Bull. Los Angel. Neuro. Soc. 27 , 169–172 (1962).
Akelaitis, A. J. A study of gnosis, praxis and language following section of the corpus callosum and anterior commissure. J. Neurosurg. 1 , 94–102 (1944).
Myers, R. E. Function of the corpus callosum in interocular transfer. Brain 79 , 358–363 (1956).
Myers, R. E. & Sperry, R. W. Interhemispheric communication through the corpus callosum: mnemonic carry-over between the hemispheres. Arch. Neurol. Psychiatry 80 , 298–303 (1958).
CAS Google Scholar
Gazzaniga, M. S. Split brain research: a personal history. Cornell Univ. Alumni Q. 45 , 2–12 (1982).
Lettvin, J. Y. 1981 Nobel prize for physiology or medicine. Science 214 , 517–520 (1981).
Gazzaniga, M. S. Cerebral specialization and interhemispheric communication: does the corpus callosum enable the human condition? Brain 123 , 1293–1326 (2000).
PubMed Google Scholar
Zaidel, E. in Handbook of Neuropsychology Vol. 4 (eds Boller, F. & Grafman, J.) 115–150 (Elsevier, Amsterdam, 1991).
Funnell, M. G., Corballis, P. M. & Gazzaniga, M. S. Handbook of Neuropsychology 2nd Edn Vol. 1 (eds Boller, F. & Grafman, J.) 103–120 (Elsevier, Amsterdam, 2000).
Milner, B. in Interhemispheric Relations and Cerebral Dominance (ed. Mountcastle, V. B.) 177–198 (Johns Hopkins Press, Baltimore, Maryland, 1962).
Zaidel, E. & Peters, A. M. Phonological encoding and ideographic reading by the disconnected right hemisphere: two case studies. Brain and Language 14 , 205–234 (1981).
Zaidel, E. in The Dual Brain (eds Benson, D. F. & Zaidel, E.) 205–231 (Guildford, New York, 1985).
Baynes, K., Eliassen, J. C., Lutsep, H. L & Gazzaniga, M. S. Modular organization of cognitive systems masked by interhemispheric integration. Science 280 , 902–905 (1998).
Nebes, R. Superiority of the minor hemisphere in commissurotomized man on a test of figural unification. Brain 95 , 633–638 (1972).
Nebes, R. Perception of spatial relationships by the right and left hemispheres of a commissurotomized man. Neuropsychologia 11 , 285–289 (1973).
Forster, B. A., Corballis, P. M. & Corballis, M. C. Effect of luminance on successiveness discrimination in the absence of the corpus callosum. Neuropsychologia 38 , 441–450 (2000).
Corballis, M. C. & Sergent, J. Imagery in a commissurotomized patient. Neuropsychologia 26 , 13–26 (1988).
Corballis, P. M., Funnell, M. G. & Gazzaniga, M. S. A dissociation between spatial and identity matching in callosotomy patients. Neuroreport 10 , 2183–2187 (1999).
Funnell, M. G., Corballis, P. M. & Gazzaniga, M. S. A deficit in perceptual matching in the left hemisphere of a callosotomy patient. Neuropsychologia 37 , 1143–1154 (1999).
Corballis, P. M., Fendrich, R., Shapley, R. & Gazzaniga, M. S. Illusory contours and amodal completion: evidence for a functional dissociation in callosotomy patients. J. Cogn. Neurosci. 11 , 459–466 (1999).
Luck, S. J., Hillyard, S. A., Mangun, G. R. & Gazzaniga, M. S. Independent hemispheric attentional systems mediate visual search in split-brain patients. Nature 342 , 543–545 (1989).
Lambert, A. J. Interhemispheric interaction in the split-brain. Neuropsychologia 29 , 941–948 (1991).
Corballis, M. C. Split decisions: problems in the interpretation of results from commissurotomized subjects. Behav. Brain Res. 64 , 163–172 (1994).
Levy, J. & Trevarthen, C. Metacontrol of hemispheric function in human split-brain patients. J. Exp. Psychol. Hum. Percept. Perform. 2 , 299–312 (1976).
Holtzman, J. D. & Gazzaniga, M. S. Dual task interactions due exclusively to limits in processing resources. Science 218 , 1325–1327 (1982).
Weissman, D. H. & Banich, M. T. The cerebral hemispheres cooperate to perform complex but not simple tasks. Neuropsychology 14 , 41–59 (2000).
Belger, A. & Banich, M. T. Costs and benefits of integrating information between the cerebral hemispheres: a computational perspective. Neuropsychology 12 , 380–398 (1998).
Banich, M. T. & Belger, A. Interhemispheric interaction: how do the hemispheres divide and conquer a task? Cortex 26 , 77–94 (1990).
Gordon, H. W., Bogen, J. E. & Sperry, R. W. Absence of deconnexion syndrome in two patients with partial section of the neocommissures. Brain 94 , 327–336 (1971).
Gazzaniga, M. S. & Freedman, H. Observations on visual processes after posterior callosal section. Neurology 23 , 1126–1130 (1973).
Risse, G. L., Gates, J., Lund, G., Maxwell, R. & Rubens, A. Interhemispheric transfer in patients with incomplete section of the corpus-callosum. Anatomic verification with magnetic resonance imaging. Arch. Neurol. 46 , 437–443 (1989).
Gazzaniga, M. S. The split brain in man. Sci. Am. 217 , 24–29 (1967).
Corballis, M. C. Visual integration in the split brain. Neuropsychologia 33 , 937–959 (1995).
Baynes, K. Language and reading in the right hemisphere: highways or byways of the brain? J. Cogn. Neurosci. 2 , 159–179 (1990).
Gazzaniga, M. S. Interhemispheric communication of visual learning. Neuropsychologia 4 , 183–189 (1966).
Seymour, S. A., Reuter-Lorenz, P. A. & Gazzaniga, M. S. The disconnection syndrome: basic findings reaffirmed. Brain 117 , 105–115 (1994).
Gazzaniga, M. S., Bogen, J. E. & Sperry, R. W. Observations on visual perception after disconnexion of the cerebral hemispheres in man. Brain 88 , 221–236 (1965).
Funnell, M. G., Corballis, P. M. & Gazzaniga, M. S. Cortical and subcortical interhemispheric interactions following partial and complete callosotomy. Arch. Neurol. 57 , 185–189 (2000).
Funnell, M. G., Corballis, P. M. & Gazzaniga, M. S. Insights into functional specificity of the human corpus callosum. Brain 123 , 920–926 (2000).
Fabri, M. et al. Posterior corpus callosum and interhemispheric transfer of somatosensory information: an fMRI and neuropsychological study of a partially callosotomized patient. J. Cogn. Neurosci. 13 , 1071–1079 (2001).
Ihori, N., Kawamura, M., Fukuzawa, K. & Kamaki, M. Somesthetic disconnection syndromes in patients with callosal lesions. Eur. Neurol. 44 , 65–71 (2000).
Arguin, M. et al. Divided visuo-spatial attention systems with total and anterior callosotomy. Neuropsychologia 15 , 295–302 (2000).
Basser, P. J. & Jones, D. K. Diffusion-tensor MRI: theory, experimental design and data analysis - a technical review. NMR Biomed. 15 , 456–467 (2002).
Basser, P. J., Mattiello, J. & LeBihan, D. Estimation of the effective self-diffusion tensor from the NMR spin echo. J. Magn. Reson. B 103 , 247–254 (1994).
Basser, P. J., Mattiello, J. & LeBihan, D. MR diffusion tensor spectroscopy and imaging. Biophys. J. 66 , 259–267 (1994).
CAS PubMed PubMed Central Google Scholar
Le Bihan, D. Looking into the functional architecture of the brain with diffusion MRI. Nature Rev. Neurosci. 4 , 469–480 (2003).
Sundgren, P. C. et al. Diffusion tensor imaging of the brain: review of clinical applications. Neuroradiology 46 , 339–350 (2004).
Chepuri, N. B. et al. Diffusion anisotropy in the corpus callosum. Am. J. Neuroradiol. 23 , 803–808 (2002).
Baird, A. A., Colvin, M. K., Van Horn, J. D., Inati, S. & Gazzaniga, M. S. Functional connectivity: integrating behavioral, DTI and fMRI data sets. J. Cogn. Neurosci. 17 , 687–693 (2005).
Warrington, E. K. & Taylor, A. M. The contribution of the right parietal lobe to object recognition. Cortex 9 , 152–164 (1973).
Humphreys, G. W., Price, C. J. & Riddoch, M. J. From objects to names: a cognitive neuroscience approach. Psychol. Res. 62 , 118–130 (1999).
Colvin, M. K., Funnell, M. G., Hahn, B. & Gazzaniga, M. S. Identifying functional channels in the corpus callosum: correlating interhemispheric transfer time with white matter organization. Poster presented at the annual meeting of the Society for Neuroscience, San Diego, California, 2004. J. Cogn. Neurosci. 139 (suppl. 5), (2005).
Aboitiz, F. & Montiel, J. One hundred million years of interhemispheric communication: the history of the corpus callosum. Braz. J. Med. Biol. Res. 36 , 409–420 (2003).
LaMantia, A. S. & Rakic, P. Cytological and quantitative characteristics of four cerebral commissures in the rhesus monkey. J. Comp. Neurol. 291 , 520–537 (1990).
Banich, M. T. The missing link: the role of interhemispheric interaction in attentional processing. Brain Cogn. 36 , 128–157 (1998).
Cabeza, R. Hemispheric asymmetry reduction in older adults: the HAROLD model. Psychol. Aging 17 , 85–100 (2002).
Colvin, M. K., Wig, G. S., Kelley, W. M., Grafton, S. T. & Gazzaniga, M. S. Callosal organization predicts the level and effect of right frontal activity during verbal encoding on subsequent memory in healthy young adults. Soc. Neurosci. Abstr. 204.4 (2005).
Colvin, M. K. Individual differences in callosal organization: relationship to interhemispheric communication and hemispheric asymmetries. Diss. Abstr. (in the press).
Leslie, A. M. & Keeble, S. Do six-month-old infants perceive causality? Cognition 25 , 265–288 (1987).
Michotte, A. The Perception of Causality (Basic Books, New York, USA, 1963) (Translated from original, published 1946).
Roser, M. E., Fugelsang, J. A., Dunbar, K. N., Corballis, P. M. & Gazzaniga, M. S. Dissociating causal perception and causal inference in the brain. Neuropsychology (in the press).
Fugelsang, J. A., Roser, M. E., Corballis, P. M., Gazzaniga, M. S. & Dunbar, K. N. Brain mechanisms underlying perceptual causality. Cogn. Brain Res. (in the press).
Turk, D. J., Heatherton, T. F., Macrae, C. N., Kelley, W. M. & Gazzaniga, M. S. Out of contact, out of mind: the distributed nature of self. Ann. NY Acad. Sci. 1001 , 65–78 (2003).
Gazzaniga, M. S. One brain — two minds? Am. Sci. 60 , 311–317 (1972).
Gazzaniga, M. S. & Smylie, C. S. Facial recognition and brain asymmetries: clues to underlying mechanisms. Ann. Neurol. 13 , 536–540 (1983).
DeRenzi, E. Prosopagnosia in two patients with CT scan evidence of damage confined to the right-hemisphere. Neuropsychologia 24 , 385–389 (1986).
Landis, T., Cummings, J. L., Christen, L., Bogen, J. E. & Imhof, H. G. Are unilateral right posterior cerebral lesions sufficient to cause prosopagnosia? Clinical and radiological findings in six additional patients. Cortex 22 , 243–252 (1986).
Michel, F., Poncet, M. & Signoret, J. L. Les lesions responsables de la prosopagnosie sont-elles toujours bilateral. Rev. Neurol. (Paris) 145 , 764–770 (1989) (in French).
Wada, Y. & Yamamoto, T. Selective impairment of facial recognition due to a haematoma restricted to the right fusiform and lateral occipital region. J. Neurol. Neurosurg. Psychiatry 71 , 254–257 (2001).
Whiteley, A. M. & Warrington, E. K. Prosopagnosia: a clinical, psychological, and anatomical study of three patients. J. Neurol. Neurosurg. Psychiatry 40 , 395–403 (1977).
Keenan, J. P., Nelson, A., O'Connor, M. & Pascual-Leone, A. Neurology: self-recognition and the right hemisphere. Nature 409 , 305 (2001).
Keenan, J. P. et al. Left hand advantage in a self-face recognition task. Neuropsychologia 37 , 1421–1425 (1999).
Keenan, J. P., Ganis, G, Freund, S. & Pascual-Leone, A. Self-face identification is increased with left hand responses. Laterality 5 , 259–268 (2000).
Conway, M. A. et al. A positron emission tomography (PET) study of autobiographical memory retrieval. Memory 7 , 679–702 (1999).
Conway, M. A. & Pleydell-Pearch, C. W. The construction of autobiographical memories in the self-memory system. Psychol. Rev. 107 , 261–288 (2000).
Kircher, T. T. et al. The neural correlates of intentional and incidental self processing. Neuropsychologia 40 , 683–692 (2002).
Maguire, E. A. & Mummery, C. J. Differential modulation of a common memory retrieval network revealed by positron emission tomography. Hippocampus 9 , 54–61 (1999).
Turk, D. J. Mike or me? Self-recognition in a split-brain patient. Nature Neurosci. 5 , 841–842 (2002).
Cooney, J. W. & Gazzaniga, M. S. Neurologic disorders and the structure of human consciousness. Trends Cogn. Sci. 7 , 161–164 (2003).
Erikson, T. C. Spread of epileptic discharge. Arch. Neurol. Psychiatry 43 , 429–452 (1940).
Gazzaniga, M. S., Bogen, J. E. & Sperry, R. W. Some functional effects of sectioning the cerebral commissures in man. Proc. Natl Acad. Sci. USA 48 , 1765–1769 (1962).
Gazzaniga, M. S. Effects of commissurotomy on a preoperatively learned visual discrimination. Exp. Neurol. 8 , 14–19 (1963).
Gazzaniga, M. S., Bogen, J. E. & Sperry, R. W. Dyspraxia following division of cerebral commissures. Arch. Neurol. 16 , 606–612 (1967).
Bogen, J. E. & Gazzaniga, M. S. Cerebral commissurotomy in man — minor hemisphere dominance for certain visuospatial functions. J. Neurosurg. 23 , 394–399 (1965).
Gazzaniga, M. S. & LeDoux, J. The Integrated Mind (Plenum, New York, USA, 1978).
Corballis, P. M. Visuospatial processing and the right-hemisphere interpreter. Brain Cogn. 53 , 171–176 (2003).
Download references
Acknowledgements
This research was supported by National Institutes of Health grants to the author. It was also supported by a graduate reseach fellowship from the National Science Foundation to M. Colvin. I would like to thank my collaborators, M. Colvin, M. Funnell, M. Roser and D. Turk, for their scientific input as well as their assistance in reviewing this paper. I would also like to thank R. Townsend for her editorial assistance.
Author information
Authors and affiliations.
the Center for Cognitive Neuroscience, 6162 Moore Hall, Dartmouth College, Hanover, 03755-3547, New Hampshire, USA
Michael S. Gazzaniga
You can also search for this author in PubMed Google Scholar
Ethics declarations
Competing interests.
The author declares no competing financial interests.
Related links
Further information.
Center for Cognitive Neuroscience
Rights and permissions
Reprints and permissions
About this article
Cite this article.
Gazzaniga, M. Forty-five years of split-brain research and still going strong. Nat Rev Neurosci 6 , 653–659 (2005). https://doi.org/10.1038/nrn1723
Download citation
Issue Date : 01 August 2005
DOI : https://doi.org/10.1038/nrn1723
Share this article
Anyone you share the following link with will be able to read this content:
Sorry, a shareable link is not currently available for this article.
Provided by the Springer Nature SharedIt content-sharing initiative
This article is cited by
- Xiaomeng Su
- Nina Skjæret-Maroni
Journal of NeuroEngineering and Rehabilitation (2024)
- Zhaonan Chen
Nature Communications (2023)
Impact of corpus callosum integrity on functional interhemispheric connectivity and cognition in healthy subjects
- Michele Porcu
- Luigi Cocco
Brain Imaging and Behavior (2023)
Impaired neurovascular coupling and cognitive deficits in anti-N-methyl-D-aspartate receptor encephalitis
- Yuanyuan Guo
- Yanghua Tian
Brain Imaging and Behavior (2022)
- Elizabeth Jefferies
Brain Structure and Function (2022)
Quick links
- Explore articles by subject
- Guide to authors
- Editorial policies
Sign up for the Nature Briefing newsletter — what matters in science, free to your inbox daily.


Keynote webinar | Spotlight on medication adherence
LIVE: Thursday 27th June 2024, 18:00-19:30 (CEST) Join our expert panel to discover why you need to understand the drivers of non-adherence in your patients, and how you can optimize medication adherence in your clinics to drastically improve patient outcomes.
Springer Medicine

Open Access 01-06-2020 | Epilepsy | Review
Split-Brain: What We Know Now and Why This is Important for Understanding Consciousness
Authors: Edward H. F. de Haan, Paul M. Corballis, Steven A. Hillyard, Carlo A. Marzi, Anil Seth, Victor A. F. Lamme, Lukas Volz, Mara Fabri, Elizabeth Schechter, Tim Bayne, Michael Corballis, Yair Pinto
Published in: Neuropsychology Review | Issue 2/2020
Please log in to get access to this content
Other articles of this issue 2/2020, a meta-analysis of neuropsychological predictors of outcome following stroke and other non-traumatic acquired brain injuries in adults, fuel for thought a systematic review of neuroimaging studies into glucose enhancement of cognitive performance, cognition-oriented treatments for older adults: a systematic overview of systematic reviews, olfaction is a marker of severity but not diagnosis in anorexia nervosa: a systematic review and meta-analysis, the effects of cognitive training on brain network activity and connectivity in aging and neurodegenerative diseases: a systematic review.
- Medical Journals
- Webcasts & Webinars
- CME & eLearning
- Newsletters
- ESMO Congress 2023
- 2023 ERS Congress
- ESC Congress 2023
- Advances in Alzheimer’s
- About Springer Medicine
- Diabetology
- Endocrinology
- Gastroenterology
- Geriatrics and Gerontology
- Gynecology and Obstetrics
- Infectious Disease
- Internal Medicine
- Respiratory Medicine
- Rheumatology
- Search Menu
- Sign in through your institution
- CNS Injury and Stroke
- Epilepsy and Sleep
- Movement Disorders
- Multiple Sclerosis/Neuroinflammation
- Neuro-oncology
- Neurodegeneration - Cellular & Molecular
- Neuromuscular Disease
- Neuropsychiatry
- Pain and Headache
- Advance articles
- Editor's Choice
- Author Guidelines
- Submission Site
- Why publish with this journal?
- Open Access
- About Brain
- Editorial Board
- Advertising and Corporate Services
- Journals Career Network
- Self-Archiving Policy
- Dispatch Dates
- Terms and Conditions
- Journals on Oxford Academic
- Books on Oxford Academic

Article Contents
Introduction, patients and methods, acknowledgements.
- < Previous
Split brain: divided perception but undivided consciousness
- Article contents
- Figures & tables
- Supplementary Data
Yair Pinto, David A. Neville, Marte Otten, Paul M. Corballis, Victor A. F. Lamme, Edward H. F de Haan, Nicoletta Foschi, Mara Fabri, Split brain: divided perception but undivided consciousness, Brain , Volume 140, Issue 5, May 2017, Pages 1231–1237, https://doi.org/10.1093/brain/aww358
- Permissions Icon Permissions
In extensive studies with two split-brain patients we replicate the standard finding that stimuli cannot be compared across visual half-fields, indicating that each hemisphere processes information independently of the other. Yet, crucially, we show that the canonical textbook findings that a split-brain patient can only respond to stimuli in the left visual half-field with the left hand, and to stimuli in the right visual half-field with the right hand and verbally, are not universally true. Across a wide variety of tasks, split-brain patients with a complete and radiologically confirmed transection of the corpus callosum showed full awareness of presence, and well above chance-level recognition of location, orientation and identity of stimuli throughout the entire visual field, irrespective of response type (left hand, right hand, or verbally). Crucially, we used confidence ratings to assess conscious awareness. This revealed that also on high confidence trials, indicative of conscious perception, response type did not affect performance. These findings suggest that severing the cortical connections between hemispheres splits visual perception, but does not create two independent conscious perceivers within one brain.

A depiction of the traditional view of the split brain syndrome ( top ) versus what we actually found in two split-brain patients across a wide variety of tasks ( bottom ). The canonical idea of split-brain patients is that they cannot compare stimuli across visual half-fields ( left ), because visual processing is not integrated across hemispheres. This is what we found as well. However, another key element of the traditional view is that split-brain patients can only respond accurately to stimuli in the left visual field with their left hand and to stimuli in the right visual field with their right hand and verbally. This is not what we found. Across a wide variety of tasks, we observed that split-brain patients could reliably indicate presence, location, orientation and identity of stimuli throughout the entire visual field regardless of how they responded.
Strikingly, although this clinical observation features in many textbooks ( Gazzaniga et al. , 1998 ; Gray, 2002 ) the reported data are never quantitative. For three reasons it is important to explicitly map out how often ‘blindness’ to the left visual field is indicated by verbal/right hand responses and unawareness to the right visual field is indicated by left hand responses. First, the number of split-brain patients is now rapidly decreasing, and it will soon be impossible to study this phenomenon. Second, there is some doubt about how clear-cut the textbook findings are. In one of the seminal publications on this topic, Sperry (1968) reports that split-brain patients seem blind to the left visual field when responding with the right hand and vice versa. However, in the last paragraph (p. 733), Sperry notes: ‘Although the general picture has continued to hold up in the main as described [… .] striking modifications and even outright exceptions can be found among the small group of patients examined to date’. Moreover, Levy et al. (1972) investigated perception of chimeric faces in five split-brain patients. Although not the focus of their research, they observed that all patients were better at matching a face to a sample when the face was presented in the left visual field, regardless of whether they responded with the left or the right hand (p. 65). Finally, note that there are multiple examples in the literature suggesting some kind of interhemispheric integration of information ( Corballis and Trudel, 1993 ; Corballis, 1995 ; Corballis and Corballis, 2001 ; Savazzi and Marzi, 2004 ; Savazzi et al. , 2007 ). This, like Sperry’s (1968) closing remark, casts doubt on the precise nature of the split-brain phenomenon.
Third, the status of split-brain patients may have important consequences for current dominant theories of consciousness. Congruent with the canonical view of split-brain patients, both the Global Workspace theory ( Baars, 1988 , 2005 ; Dehaene and Naccache, 2001 ) and the Information Integration theory ( Tononi, 2004 , 2005 ; Tononi and Koch, 2015 ) imply that without massive interhemispheric communication two independent conscious systems appear. If the canonical view cannot be quantitatively replicated, and evidence for conscious unity in the split-brain syndrome is found, both theories may require substantial modifications.
In our current studies we reproduced the classic finding that split-brain patients are unable to integrate visual information across the two visual half-fields. However, we also investigated systematically to what extent performance depends on where a stimulus appears. For various tasks and stimuli we studied whether there is a response type × visual field interaction: can split-brain patients only respond to stimuli in the left visual field with the left hand, and to stimuli in the right visual field with the right hand or verbally?
Patients were tested across several years, during their routine neurological control visits. For Experiment 1 we tested Patients DDC and DDV, for Experiments 2–5 we tested Patient DDC. Both patients underwent a full callosotomy to relieve epileptic seizures. Crucially, for current purposes, in Patient DDC the complete corpus callosum and most of the anterior commissure was cut, and in Patient DDV the complete corpus callosum was removed. We selected Patient DDC for the extensive follow-up testing since his ‘split’ is the most severe. Note that other than the removal of the corpus callosum, both patients had no brain damage, and fell within the normal IQ range. See Supplementary material and Pizzini et al. (2010) and Corballis et al. (2010) for detailed descriptions of these patients. In all experiments the patient(s) responded with three response types (response conditions were blocked), verbal, right hand or left hand, except for Experiment 2A, where the patient only responded verbally; and Experiments 2C and 4A where only left and right hand responses and no verbal responses were given. The experimenter (who could not see the test stimuli) mouse-clicked on the response box or location indicated by the patient. In the case of verbal position indication, the mouse was moved by the experimenter (not having seen the stimulus) on the instructions of the patient until the desired position was obtained.
In Experiment 1 both patients performed a combined detection/localization task. Either nothing appeared (50% of trials) or a red solid circle, on a grey background (see Supplementary material for all stimulus details), appeared for 120 ms anywhere in the visual field. Each trial the patient indicated whether a stimulus had appeared, and if so where.
In Experiment 2A, Patient DDC indicated whether two rectangles had the same orientation. In Experiment 2B he reported if two simple shapes were the same, and in Experiment 2C he indicated if two pictures were equal. In all experiments the test stimuli appeared for 120 ms. The stimuli appeared (i) both in left visual field; (ii) both in right visual field; or (iii) they appeared around fixation with one stimulus in left visual field and on in right visual field. In Experiment 3A a picture was presented for 120 ms in the left or right visual field, after which Patient DDC selected the correct verbal label matching the picture. Experiment 3B was identical to 3A, but instead of selecting a verbal label, Patient DDC selected from two pictures which image he had just seen.
In Experiment 4A either nothing appeared, or a simple shape (square, circle or triangle) appeared for 100 ms in the left or right visual field. Patient DDC indicated if something had appeared, and if so what. In Experiment 4B two rectangles were successively presented, the first of which appeared for 120 ms, in the left or right visual field. Patient DDC indicated whether both rectangles had the same orientation, and if not, how large the orientation difference was. In both experiments, after each trial, Patient DDC indicated confidence in his judgement (Experiment 4A on a scale from 1 to 4, Experiment 4B on a scale from 1 to 4). Experiment 5 was similar to Experiment 1, except after each trial Patient DDC indicated confidence in his presence and location judgement (on a scale from 1 to 5). Moreover, stimuli were bright green on a red background, or dim green on a red background. In the latter case stimuli and background were equiluminant (as determined by an objective measurement).
An overview of the results of Experiment 1. Both split-brain patients, Patients DDC and DDV, accurately indicated presence and location (distance error is in degrees of visual angle) of stimuli appearing throughout the entire visual field, regardless of response type (verbally, left hand or right hand). These findings challenge the canonical view that split-brain patients can only respond correctly to the left visual field with the left hand and vice versa.
In Experiment 1 ( Fig. 2 ), we explored to what extent Patients DDV and DDC can detect stimuli across the entire visual field using three response conditions: left hand, right hand, and verbally. Subjects were shown red circles in various locations of the visual field (50% of trials no stimulus was presented), and had to detect presence or absence either verbally or by indicating yes/no with either hand. Subsequently, for seen stimuli, they had to indicate the location of the stimulus. Both patients responded (nearly) perfectly in indicating presence of the stimulus (Patient DDV, hits: 100%, false alarms: 0%; Patient DDC, hits: 97.5%, false alarms: 7.7%), and were highly accurate in indicating location of the stimulus (average distance between pointed location and actual location: Patient DDV: 2.8°, Patient DDC: 4.5°). While presence and location performance was highly significantly above chance (all P < 0.001), the response type × visual half-field interaction did not approach significance in either patient or task (all P > 0.5).

An overview of the results of Experiments 2 and 3. Patient DDC was not able to compare stimuli across visual half-fields, although he was able to do so within one visual half-field (Experiment 2A–C). Moreover, he was better at labelling stimuli in the right visual field (Experiment 3A) and better at matching stimuli in the left visual field (Experiment 3B). Crucially, although visual information remained unintegrated across visual half-fields, there was still no response type × visual field interaction.
We found further evidence that visual information is not shared between hemispheres in Experiment 3 ( Fig. 3 ). Here we observed that Patient DDC was better at selecting the correct verbal label for an image when it had appeared in the right visual field than when it had appeared in the left visual field (Experiment 3A, left visual field: 73.4%, right visual field: 92.1%, left visual field versus right visual field: P < 0.001). Yet, he was better at matching a stimulus to sample for items in left visual field, replicating earlier split-brain findings ( Funnell et al. , 1999 ) (Experiment 3B, left visual field: 95.5%, right visual field: 73%, left visual field versus right visual field: P < 0.001). Note that, despite the seeming lack of transfer of visual information, we still observed no response type × visual field interaction in Experiments 2 and 3 (all P > 0.12). Thus, for instance, Patient DDC was better at matching to sample of stimuli in the left visual field even when he responded with the right hand. This suggests that processing of visual stimuli remains within each individual hemisphere, each with its own relative performances in various tasks, yet control over the report of the outcomes of this processing is undivided.

Across three experiments Patient DDC performed better on high confidence than on low confidence trials, suggesting accurate metacognition. Moreover, also for high confidence trials we observed no response type × visual field interaction, suggesting that unity in responding was based on conscious perception, not on blindsight-like processes.
First, Patient DDC was tested on two visual matching experiments (shape and orientation). Second, he performed a detection and localization task of simple stimuli as in the first experiment (with the addition that the stimuli were presented equiluminantly with the background or with a large luminance difference). Patient DDC performed nearly flawlessly in detecting objects in Experiment 5 (no false alarms and two misses in 167 trials). This ceiling effect precluded meaningful metacognitive assessment of this aspect of the task. However, in the other two experiments and the localization of objects in Experiment 5, performance did not show a ceiling or floor effect, allowing us to investigate metacognitive abilities in these cases. This revealed that in all three experiments Patient DDC’s performance was better on high than on low confidence trials. All trials: Experiment 4A, left visual field: 88.7%, right visual field: 43%; Experiment 4B, left visual field: 82.8% right visual field: 63.4%; Experiment 5, left visual field: accuracy: 100%, distance error: 3.27°, right visual field: accuracy: 98.3%, distance error: 2.33°. High confidence trials: Experiment 4A, left visual field: 100%, right visual field: 62.5%; Experiment 4B, left visual field: 95.9% right visual field: 84.6%; Experiment 5, left visual field: accuracy: 100%, distance error: 2.63°, right visual field: accuracy: 98.2%, distance error: 1.84°; all P < 0.005). Further, we found a robust Goodman and Kruskal’s γ correlation ( Goodman and Kruskal, 1954 ) between confidence and performance in all cases (Experiment 4A, γ = 0.527, P < 0.001; Experiment 4B, γ = 0.316, P = 0.003; Experiment 5, γ = −0.227, P = 0.02. There were no differences between γ correlations in left visual field and right visual field, all P > 0.09). Both analyses indicate that Patient DDC possessed accurate metacognition. Crucially, on high confidence trials we still found no response type × visual field interaction (all P > 0.63). This indicates that Patient DDC’s performance is not rooted in unconscious processes: his correct answers are based on conscious awareness and decisions. Note further that in the detection and localization task, luminance difference did not affect results (all P > 0.8), indicating that our findings are not due to overly strong stimulation, or stray-light leaking over to the other visual half-field.
In addition to these five experiments we obtained phenomenal reports from both split-brain patients (see Supplementary material for an extensive description). Both patients indicated that they saw their entire visual field (so not just the visual field to the left or right of fixation). Further, they indicated that they felt, and were in control of their entire body. Finally, they reported that their conscious unity was unchanged since the operation (i.e. no other conscious agent seemed to be present in their brain/body). These phenomenal reports are congruent with earlier reports of split-brain patients, which documented that split-brain patients feel normal and behave normally in social situations ( Bogen et al. , 1965 ; Sperry, 1968 ).
In conclusion, with two patients, and across a wide variety of tasks we have shown that severing the cortical connections between the two hemispheres does not seem to lead to two independent conscious agents within one brain. Instead, we observed that patients without a corpus callosum were able to respond accurately to stimuli appearing anywhere in the visual field, regardless of whether they responded verbally, with the left or the right hand—despite not being able to compare stimuli between visual half-fields, and despite finding separate levels of performance in each visual half-field for labelling or matching stimuli. This raises the intriguing possibility that even without massive communication between the cerebral hemispheres, and thus increased modularity, unity in consciousness and responding is largely preserved.
This preserved unity of consciousness may be especially challenging for the two currently most dominant theories of consciousness, the Global Workspace theory ( Baars, 1988 , 2005 ; Dehaene and Naccache, 2001 ) and the Integration Information theory ( Tononi, 2004 , 2005 ; Tononi and Koch, 2015 ). A core assumption of the Global Workspace theory is that cortical broadcasting of selected information by the ‘global workspace’ leads to consciousness. Thus severing of the corpus callosum, which prevents broadcasting of information across hemispheres, seems to exclude the emergence of one global workspace for both hemispheres. Rather, it seems that without a corpus callosum either two independent global workspaces emerge, or only one hemisphere will have a global workspace, while the other does not. In either case, an integrated global workspace, and thus preserved conscious unity, seems to be difficult to fit into this framework.
Also for Integration Information theory, conscious unity in the split-brain syndrome seems to be challenging. According to the Integration Information theory the richness of integration of information (called φ, defined by how much information is represented, and how integrated the information is) determines the level of consciousness. Moreover, only if the combined φ of two subsystems is larger than the φ per system, then the two subsystems combine to form one conscious entity. After removal of the corpus callosum, which all but eliminates communication between the cerebral hemispheres, integration of information is larger within each hemisphere than between hemispheres. Thus, according to the Integration Information theory, in the split-brain syndrome φ per hemisphere is larger than the combined φ, thus leading to two independent conscious systems rather than one conscious agent ( Tononi, 2005 ).
It thus seems that the current results provide a challenge for the Global Workspace and the Integrated Information theory of consciousness. However, the current results may fit well with the local recurrent processing theory of consciousness ( Lamme and Roelfsema, 2000 ; Lamme, 2006 ; Block, 2007 ). This theory claims that local recurrent interactions between neural areas (for example between V1 and V5 in the visual system) are enough to create consciousness, even if these interactions are not part of a larger integrated network, and do not project their outcomes to a central processing unit. Thus, according to this theory, even in healthy subjects, relatively isolated processing in one hemisphere can lead to normal visual experiences. Therefore, the local recurrent processing theory suggests that consciousness in split-brain patients may be similar to consciousness in healthy subjects (and thus equally unified).
How should these results be compared to our classic text-book knowledge of the split-brain phenomenon? It is unlikely that our results can be explained by the anterior and posterior commissure still being (somewhat) intact, as this was also the case for many of the previously tested patients, and this did not seem to play an important role then ( Gazzaniga et al. , 1985 ; Seymour et al. , 1994 ; Gazzaniga, 2005 ).
Another possible explanation to consider is that the current findings were caused by cross-cueing (one hemisphere informing the other hemisphere with behavioural tricks, such as touching the left hand with the right hand). We deem this explanation implausible for four reasons. First, cross-cueing is thought to only allow the transfer of one bit of information ( Baynes et al. , 1995 ). Yet, both patients could localize stimuli throughout the entire visual field irrespective of response mode (Experiments 1 and 5), and localizing a stimulus requires more than one bit of information. Second, visual capabilities differed per hemifield (Experiment 3: better matching for stimuli in left visual field, better labelling of stimuli in right visual field) and comparison of stimuli over hemifields was not possible (Experiment 2). This suggests that transfer of visual information did not occur. Yet, in these same experiments response type did not affect performance, suggesting that unity in control was not driven by any form of transfer of visual information. Third, we explicitly set up the experiments to prevent cross-cueing (e.g. hands were not allowed to touch each other, or the other half of the body). Moreover, we did not observe any indications of cross-cueing occurring. Fourth, as cross-cueing is a slow process, ipsilateral responses driven by cross-cueing should be considerably slower than contralateral responses. Yet, in one experiment where Patient DDC indicated, as quickly as possible, the colour of a circle appearing shortly to the left or the right of fixation, average ipsilateral and contralateral responses were almost equally fast, and equally accurate (ipsilateral reaction times: 1229 ms, ipsilateral accuracy: 88.4%; contralateral reaction times: 1307 ms, contralateral accuracy: 97%; No significant difference between ipsilateral and contralateral reaction times: P = 0.13; or between ipsilateral and contralateral accuracy: P = 0.55, see Supplementary material for details).
Finally, a possibility is that we observed the current results because we tested these patients well after their surgical removal of the corpus callosum (Patient DDC and Patient DDV were operated on at ages 19 and 22 years, and were tested 10–16 and 17–23 years after the operation, respectively). This would raise the interesting possibility that the original split brain phenomenon is transient, and that patients somehow develop mechanisms or even structural connections to re-integrate information across the hemispheres, particularly when operated at early adulthood. Even then, it remains the case that these patients’ minds have a curious property: somehow, their perception seems split, each hemisphere processing visual information independently, and at the best of their individual—yet different—abilities. When it comes to reporting this information to the outside world, however, the outcomes of the perceptual processes are unified in consciousness, verbalization and control of the body. This ‘split phenomenality’ combined with ‘unity of consciousness’ is difficult to grasp introspectively, and surely warrants further study, in a group of patients of which very few remain today.
We thank Gabriella Venanzi for scheduling the patients’ exams. We also thank D.D.C. and D.D.V. and their families for their willingness to collaborate in these studies.
This research was supported by a Marie Curie IEF grant (PIEF-GA-2011-300184) to Y.P., a Marie Curie IEF grant (PIEF-GA-2012-SOC-329134) to M.O. and by Advanced Investigator Grants by the European Research Council to V.A.F.L. and E.H.FdH.
Supplementary material
Supplementary material is available at Brain online.
Baars BJ . A cognitive theory of consciousness . New York : Cambridge University Press ; 1988 .
Google Scholar
Google Preview
Baars BJ . Global workspace theory of consciousness: toward a cognitive neuroscience of human experience . Prog Brain Res 2005 ; 150 : 45 – 53 .
Baynes K , Wessinger CM , Fendrich R , Gazzaniga MS . The emergence of the capacity to name left visual field stimuli in a callosotomy patient: implications for functional plasticity . Neuropsychologia 1995 ; 33 : 1225 – 42 .
Block N . Consciousness, accessibility, and the mesh between psychology and neuroscience . Behav Brain Sci 2007 ; 30 : 481 – 99 .
Bogen JE , Fisher E , Vogel P . Cerebral commissurotomy: a second case report . JAMA 1965 ; 194 : 1328 – 9 .
Cantalupo C , Hopkins WD . Asymmetric broca's area in great apes . Nature 2001 ; 414 : 505 .
Corballis MC . Visual integration in the split brain . Neuropsychologia 1995 ; 33 : 937 – 59 .
Corballis MC , Birse K , Paggi A , Manzoni T , Pierpaoli C , Fabri M . Mirror-image discrimination and reversal in the disconnected hemispheres . Neuropsychologia 2010 ; 48 : 1664 – 9 .
Corballis MC , Corballis PM . Interhemispheric visual matching in the split brain . Neuropsychologia 2001 ; 39 : 1395 – 400 .
Corballis MC , Trudel CI . Role of the forebrain commissures in interhemispheric integration . Neuropsychology 1993 ; 7 : 306 .
Cowey A . Cortical maps and visual perception the grindley memorial lecture . Q J Exp Psychol 1979 ; 31 : 1 – 17 .
Dehaene S , Naccache L . Towards a cognitive neuroscience of consciousness: basic evidence and a workspace framework . Cognition 2001 ; 79 : 1 – 37 .
Funnell M , Corballis P , Gazzaniga M . A deficit in perceptual matching in the left hemisphere of a callosotomy patient . Neuropsychologia 1999 ; 37 : 1143 – 54 .
Gazzaniga MS , Ivry RB , Mangun GR . The problem of consciousness . In: Cognitive neuroscience - the biology of the mind . New York, NY : W. W. Norton & Company ; 1998 . p. 542 – 3 .
Gazzaniga MS . The split brain in man . Sci Am 1967 ; 217 : 24 – 29 .
Gazzaniga MS . Forty-five years of split-brain research and still going strong . Nat Rev Neurosci 2005 ; 6 : 653 – 9 .
Gazzaniga MS . Cerebral specialization and interhemispheric communication: does the corpus callosum enable the human condition? Brain 2000 ; 123 : 1293 – 326 .
Gazzaniga MS , Bogen JE , Sperry RW . Some functional effects of sectioning the cerebral commissures in man . Proc Natl Acad Sci USA 1962 ; 48 : 1765 – 9 .
Gazzaniga MS , Holtzman JD , Deck MD , Lee BC . MRI assessment of human callosal surgery with neuropsychological correlates . Neurology 1985 ; 35 : 1763 – 6 .
Goodman LA , Kruskal WH . Measures of association for cross classifications . J Am Stat Assoc 1954 ; 49 : 732 – 64 .
Gray P . The nervous system . Psychology . New York, NY : Worth Publishers ; 2002 . p. 160 – 1 .
Innocenti GM . General organization of callosal connections in the cerebral cortex . In: Jones EG , Peters A , editors, Sensory-motor areas and aspects of cortical connectivity . New York : Plenum Springer ; 1986 . pp. 291 – 353 .
Lamme VA . Towards a true neural stance on consciousness . Trends Cogn Sci 2006 ; 10 : 494 – 501 .
Lamme VA , Roelfsema PR . The distinct modes of vision offered by feedforward and recurrent processing . Trends Neurosci 2000 ; 23 : 571 – 9 .
Lau H , Rosenthal D . Empirical support for higher-order theories of conscious awareness . Trends Cogn Sci 2011 ; 15 : 365 – 73 .
Levy J , Trevarthen C , Sperry RW . Reception of bilateral chimeric figures following hemispheric deconnexion . Brain 1972 ; 95 : 61 – 78 .
Ojemann G , Ojemann J , Lettich E , Berger M . Cortical language localization in left, dominant hemisphere: an electrical stimulation mapping investigation in 117 patients . J Neurosurg 1989 ; 71 : 316 – 26 .
Penfield W , Boldrey E . Somatic motor and sensory representation in the cerebral cortex of man as studied by electrical stimulation . Brain 1937 ; 60 : 389 .
Pizzini F , Polonara G , Mascioli G , Beltramello A , Foroni R , Paggi A et al. Diffusion tensor tracking of callosal fibers several years after callosotomy . Brain Res 2010 ; 1312 : 10 – 17 .
Rosenthal D . Consciousness and metacognition . In: Sperber D , editor. Metarepresentation: Proceedings of the tenth vancouver cognitive science conference . New York : Oxford University Press ; 2000 . p. 265 – 95 .
Sakata H , Taira M . Parietal control of hand action . Curr Opin Neurobiol 1994 ; 4 : 847 – 56 .
Savazzi S , Fabri M , Rubboli G , Paggi A , Tassinari CA , Marzi CA . Interhemispheric transfer following callosotomy in humans: role of the superior colliculus . Neuropsychologia , 2007 ; 45 : 2417 – 27 .
Savazzi S , Marzi CA . The superior colliculus subserves interhemispheric neural summation in both normals and patients with a total section or agenesis of the corpus callosum . Neuropsychologia 2004 ; 42 : 1608 – 18 .
Seymour SE , Reuter-Lorenz PA , Gazzaniga MS . The disconnection syndrome - basic findings reaffirmed . Brain 1994 ; 117 : 105 – 115 .
Sperry R . Consciousness, personal identity and the divided brain . Neuropsychologia 1984 ; 22 : 661 – 73 .
Sperry RW . Hemisphere deconnection and unity in conscious awareness . Am Psychol 1968 ; 23 : 723 .
Tononi G . Consciousness, information integration, and the brain . Prog Brain Res 2005 ; 150 , 109 – 26 .
Tononi G . An information integration theory of consciousness . BMC Neurosci 2004 ; 5 : 42 .
Tononi G , Koch C . Consciousness: here, there and everywhere? Philos Trans R Soc Lond B Biol Sci 2015 ; 370 . pii: 2014.0167 .
Vigneau M , Beaucousin V , Herve P , Duffau H , Crivello F , Houde O et al. Meta-analyzing left hemisphere language areas: phonology, semantics, and sentence processing . Neuroimage 2006 ; 30 : 1414 – 32 .
Wahl M , Lauterbach-Soon B , Hattingen E , Jung P , Singer O , Volz S et al. Human motor corpus callosum: topography, somatotopy, and link between microstructure and function . J Neurosci 2007 ; 27 : 12132 – 8 .
Wolman D . A tale of two halves . Nature , 2012 ; 483 : 260 – 3 .
- consciousness related finding
- corpus callosum
- visual fields
Supplementary data
Email alerts, citing articles via, looking for your next opportunity.
- Contact the editorial office
- Guarantors of Brain
- Recommend to your Library
Affiliations
- Online ISSN 1460-2156
- Print ISSN 0006-8950
- Copyright © 2024 Guarantors of Brain
- About Oxford Academic
- Publish journals with us
- University press partners
- What we publish
- New features
- Open access
- Institutional account management
- Rights and permissions
- Get help with access
- Accessibility
- Advertising
- Media enquiries
- Oxford University Press
- Oxford Languages
- University of Oxford
Oxford University Press is a department of the University of Oxford. It furthers the University's objective of excellence in research, scholarship, and education by publishing worldwide
- Copyright © 2024 Oxford University Press
- Cookie settings
- Cookie policy
- Privacy policy
- Legal notice
This Feature Is Available To Subscribers Only
Sign In or Create an Account
This PDF is available to Subscribers Only
For full access to this pdf, sign in to an existing account, or purchase an annual subscription.
One Head, Two Brains
How a radical epilepsy treatment in the early 20th century paved the way for modern-day understandings of perception, consciousness, and the self

In 1939, a group of 10 people between the ages of 10 and 43, all with epilepsy, traveled to the University of Rochester Medical Center, where they would become the first people to undergo a radical new surgery.
The patients were there because they all struggled with violent and uncontrollable seizures. The procedure they were about to have was untested on humans, but they were desperate—none of the standard drug therapies for seizures had worked.
Between February and May of 1939, their surgeon William Van Wagenen, Rochester’s chief of neurosurgery, opened up each patient’s skull and cut through the corpus callosum, the part of the brain that connects the left hemisphere to the right and is responsible for the transfer of information between them. It was a dramatic move: By slicing through the bundle of neurons connecting the two hemispheres, Van Wagenen was cutting the left half of the brain away from the right, halting all communication between the two.
In a paper he and a colleague published in the Journal of the American Medical Association in 1940, Van Wagenen explained his reasoning: He had developed the idea for the surgery after observing two epilepsy patients with brain tumors located in the corpus callosum. The patients had experienced frequent convulsive seizures in the early stages of their cancer, when the tumors were still relatively small masses in the brain—but as the tumors grew, they destroyed the corpus callosum, and the seizures eased up.
“In other words, as the corpus callosum was destroyed, generalized convulsive seizures became less frequent,” Van Wagenen wrote in the 1940 paper, noting that “as a rule, consciousness is not lost when the spread of the epileptic wave is not great or when it is limited to one cerebral cortex.” Based on the cases of the cancer patients—and some other clinical observations —Van Wagenen believed that destroying the corpus callosum of his patients would block the spread of the electrical impulses that lead to seizures, so that a seizure that began in the left hemisphere, for example, stayed in the left hemisphere.
The surgery worked for most of the patients : In his paper, Van Wagenen reported that seven of the 10 experienced seizures that were less frequent or less severe.
Between 1941 and 1945, Van Wagenen’s colleague, the University of Rochester psychiatrist A. J. Akelaitis, tested the patients to see if they had experienced any cognitive or behavioral changes as a result of the invasive procedure. After giving the patients a series of assessments—an I.Q. test, a memory test, motor-skills assessments, and interviews—he reported that most of the patients had the same levels of cognitive functioning after the surgery as before, and displayed no behavioral or personality changes. Though the brain hemispheres of split-brain patients had been disconnected, he wrote in a 1944 paper in the Journal of Neurosurgery , they were otherwise normal.
Or so it seemed.
When Michael Gazzaniga first learned about the Rochester patients as an undergraduate research intern in 1960, he was curious—and skeptical.
Gazzaniga’s timing was fortuitous: Roger Sperry, who headed the neuroscience lab where Gazzaniga worked at the California Institute of Technology, had begun split-brain research on cats and monkeys just a few years earlier. Sperry found that severing the corpus callosum of those animals had affected their behavior and cognitive functioning.
In one experiment with split-brain cats, for example, Sperry would cover one of the animal’s eyes and then teach it to differentiate between a triangle and a square. Once the cats learned to do that, Sperry switched the covering from one eye to the other and tested the them to see if they recalled their new knowledge. They didn’t. “The split-brain cat,” as one neurosurgeon wrote in an overview of Sperry’s work, “has to learn all over again.” As Sperry noted, this suggested that the two hemispheres were not communicating with each other, and that each was learning the task on its own.
If the Rochester patients’ left and right brains were also no longer communicating, Sperry and his colleagues believed, then they must be experiencing some sort of change, too.
The question was still bothering Gazzaniga by the time he returned to Sperry’s lab as a graduate student in 1961: What kind of change was it? Would human brains react the same way as those of the animals in Sperry’s lab?
“In monkeys,” Gazzaniga told me, “sectioning the corpus callosum led to the right hand not knowing what the left hand was doing. I wanted to know if we would see a similar result in humans.”
The researchers didn’t have to wait long to begin looking for the answer. In the summer of 1961, as Gazzaniga was preparing to return to Sperry’s lab as a graduate student, a young neurosurgeon at Caltech named Joseph Bogen approached Sperry about the opportunity to study a split-brain patient—and Sperry, who had been working exclusively with animals, seized the chance to work on his first human case.
The patient Bogen had in mind was a man in his late forties named William Jenkins, a World War II veteran who had been hit in the head with the butt of a German officer’s rifle after parachuting behind enemy lines. Jenkins’ doctors believed that this was the likely origin of the uncontrollable seizures he later developed; when he returned to the U.S. after the war and sought treatment, he discovered that no drugs worked to contain the seizures.
In 1961, as a last-ditch effort, Bogen suggested that he have split-brain surgery. Sperry assigned Gazzaniga to conduct some standard pre-operative neurological tests, and Bogen and a colleague performed the procedure in February of 1962. After a few months of post-surgery monitoring, Bogen found that the severity and frequency of Jenkins’ seizures had abated, but he still did not know if the surgery had produced other unintended consequences. So about a month after the surgery, Bogen sent Jenkins to Sperry and Gazzaniga for cognitive testing. In doing so, he kicked off a line of work that would turn the two men into pioneers of split-brain research, eventually earning Sperry a share of the Nobel Prize in 1981—and causing scientists to reconsider long-held ideas about the brain and the self.
The cognitive tests performed on the 10 original Rochester patients hadn’t tested each brain hemisphere separately; believing that this was one reason why the patients hadn’t shown any changes after surgery, Sperry and Gazzaniga decided to run tests for both the left and right sides of Jenkins’s brain.
Recommended Reading

The Personality Trait That Makes People Feel Comfortable Around You

Loving Your Job Is a Capitalist Trap

The Seven Habits That Lead to Happiness in Old Age
In one of the first split-brain studies that the pair designed, published in August 1962 in the Proceedings of the National Academy of Sciences , Gazzaniga invited Jenkins into the lab and had him stare straight ahead at a dot. As he was staring ahead, Gazzaniga flashed a picture of a square on a screen to the right of where his eyes were staring, meaning the image would be processed by Jenkins’ left brain. ( Because of the way the brain is wired , if a patient looks straight ahead, something quickly flashed to the left of his gaze will be processed by the right side of the brain, and vice versa. The brain’s hemispheres control activity mainly on the opposite side of the body—the left hemisphere controls the action of the right hand, for example, while the right hemisphere moves the left hand.)
When Gazzaniga asked Jenkins what he saw, Jenkins was able to describe the square. Then Gazzaniga tried the same thing on the other side, flashing the same image to the left of Jenkins’ gaze. When he asked Jenkins again what he saw, though, Jenkins said he saw nothing.
Intrigued, Gazzaniga pulled another image, this time of a circle, to flash on Jenkins’s right and left sides separately, as he had done with the square.
Instead of asking Jenkins to describe the object, though, he asked him to point to it. When the image was on Jenkins’ right side (left brain), he lifted his right hand (controlled by the left brain) to point to it. When the circle flashed on his left side (right brain), he lifted his left hand (controlled by the right brain) to point to it.
The fact that Jenkins was able to point to the circle with both hands told Gazzaniga that each of Jenkins’ hemispheres had processed the sight of the circle. It also meant that in the previous trial, both of Jenkins’s hemispheres had processed the square—even though Jenkins said, when his right brain processed the sight, that he saw nothing. At that point, scientists had known for about a century that language arises from the left hemisphere; given that, the researchers later reasoned, Jenkins could only talk about the square when its picture was flashed to his right eye (left brain). On the other side, even though Jenkins had seen the square, he could not speak about it.
Between 1962 and 1967, Sperry and Gazzaniga worked together to perform dozens of additional experiments with Jenkins and other split-brain patients. In one set of studies conducted in 1962 and 1963, Gazzaniga presented Jenkins with four multicolored blocks. Then, he showed Jenkins a picture of the blocks arranged in a certain order, and asked him to make the same arrangement with the blocks in front of him.
Because the right brain handles visual-motor capacity, Gazzaniga was unsurprised to see that Jenkins’ right hemisphere excelled at this task: Using his left hand, Jenkins was immediately able to arrange the blocks correctly. But when he tried to do the very same task with his right hand, he couldn’t. He failed, badly.
“It couldn’t even get the overall organization of how the blocks should be positioned, in a 2x2 square,” Gazzaniga later wrote of Jenkins’ left hemisphere in his memoir, Tales from Both Sides of the Brain . “It just as often would arrange them in a 3+1 shape.”
But more surprising was this: As the right hand kept trying to get the blocks to match up to the picture, the more capable left hand would creep over to the right hand to intervene, as if it realized how incompetent the right hand was. This occurred so frequently that Gazzaniga eventually asked Jenkins to sit on his left hand so it wouldn’t butt in.
When Gazzaniga let Jenkins use both hands to solve the problem in another trial, he again saw the two brain hemispheres at odds with one another. “One hand tried to undo the accomplishments of the other,” he wrote. “The left hand would make a move to get things correct and the right hand would undo the gain. It looked like two separate mental systems were struggling for their view of the world.”
The more information the split-brain researchers discovered, the more they wondered: If the two sides of the brain functioned so independently of each other, how do people—ordinary people and split-brain patients alike—experience a single, cohesive reality?
In a 1977 study with a 15-year-old split-brain patient from Vermont identified as P. S., Gazzaniga (then a professor at Dartmouth) and his graduate assistant Joseph LeDoux performed a visual test similar to the one Jenkins had undergone years earlier. The researchers asked P. S. to stare straight ahead at a dot, and then flashed a picture of a chicken foot to the brain’s left hemisphere and a picture of a snowy scene to the brain’s right hemisphere. Directly in front of the patient—so that he could process the sight with both hemispheres—was a series of eight other pictures. When the researchers asked him to point to the ones that went with the images he saw, P. S. pointed to the picture of a chicken head and a picture of a snow shovel.
So far, the results were as expected: Each hemisphere had led P. S. to choose an image that went along with the one that he had seen from that side moments earlier. The surprise came when the researchers asked him why he chose these two totally unrelated images.
Because the left hemisphere, which controls language, had not processed the snowy scene, they believed P. S. wouldn’t be able to verbally articulate why he chose the snow shovel. “The left brain doesn’t know why,” Gazzaniga told me. “That information is in the right hemisphere.” Neither hemisphere knew what the other had seen, and because the two sides of his brain were unable to communicate, P.S. should have been confused when Gazzaniga asked him why he had picked the two images he did.
But as Gazzaniga recalled in his memoir, P. S. didn’t skip a beat: “Oh, that’s simple,” the patient told them. “The chicken claw goes with the chicken, and you need a shovel to clean out the chicken shed.”
Here’s what happened, as the researchers later deduced: Rather leading him to simply say, “I don’t know” to Gazzaniga’s question, P.S.’s left brain concocted an answer as to why he had picked those two images. In a brief instant, the left brain took two unconnected pieces of information it had received from the environment—the two images—and told a story that drew a connection between them.
Gazzaniga went on to replicate the findings of this study many times with various co-authors: When faced with incomplete information, the left brain can fill in the blanks. Based on these findings Gazzaniga developed the theory that the left hemisphere is responsible for our sense of psychological unity—the fact that we are aware of and reflect upon what is happening at any given moment.
“It’s the part of the brain,” Gazzaniga told me, “that takes disparate points of information in and weaves them into a storyline and meaning. That it’s central gravity.”
In addition to answering questions of brain specialization, split-brain research also examined some of the ways in which the left and right hemispheres are autonomous agents. Jenkins’ left and right hands started fighting over how to arrange the blocks, for example, because the two hemispheres are—as Gazzaniga told me—“two separate minds, all in one head.”
As he further explained in Tales from Both Sides of the Brain : “The notion that there is an ‘I’ or command center in the brain was an illusion.”
Among psychologists, the idea wasn’t exactly new; figures like Sigmund Freud and William James had previously theorized about a “divided self,” with Freud arguing that the mind is divided into the ego, the superego, and the id. But split-brain research was arguably one of the first scientific demonstrations that the divided self has a real, physical basis—a demonstration that, in turn, raised new questions about the relationship between the mind and the brain.
“The demonstration that you could in effect split consciousness by splitting anatomy—by just making a tiny change in anatomy … It was one of the most remarkable results in neuroscience, with huge implications,” said Patricia Churchland, a philosopher at the University of California, San Diego, whose work focuses on the relationship between philosophy and neuroscience. “If you thought that consciousness and mental states were independent of the brain, then this should have been a real wake-up call.”
Helping to illuminate the relationship between the mind and the brain, according to the cognitive psychologist Steven Pinker, is one of split-brain research’s most important contributions to modern psychology and neuroscience. “The fact that each hemisphere supports its own coherent, conscious stream of thought highlights that consciousness is a product of brain activity,” he told me. “The notion that there is a single entity called consciousness , without components or parts, is false.”
Today’s therapies for seizures are more advanced than those of the mid-20th century, and split-brain surgery is now exceedingly rare —Michael Miller, a neuroscientist at the University of California at Santa Barbara who did graduate work with Gazzaniga, told me the last one he heard of was performed around 10 years ago. Many of the split-brain patients that Gazzaniga, Sperry, and their colleagues studied have passed away.
Though the research on split-brain patients has slowed dramatically, Miller believes that the field still has something left to offer. He’s currently working on a study currently working with a patient to answer the question: Does each hemisphere of the brain reflect on and evaluate itself in a unique way?
“We know that the two hemispheres have different strategies for thinking,” Miller told me, “and we’re curious about how that might change their reflection of themselves. Does the left hemisphere think of itself as a sad person while the right one think of itself as a happy person? We are having each hemisphere evaluate itself to find out.”
Miller’s study uses a test called the “trait-judgment task”: A trait like happy or sad flashes on a screen, and research subjects indicate whether the trait describes them. Miller has slightly modified this task for his split-brain patients—in his experiments, he flashes the trait on a screen straight in front of the subject’s gaze, so that both the left and right hemispheres process the information. Then, he quickly flashes the words “me” and “not me” to one side of the subject’s gaze—so that they’re processed only by one hemisphere—and the subject is instructed to point at the trait on the screen when Miller flashes the appropriate descriptor. (For example, if the screen reads “happy,” an unhappy left hemisphere would lead a subject to point when Miller flashes “not me” to the right side of the subject’s gaze, and to stay still when he flashes “me.”) If the subject reacts differently on each side—in this example, if the subject points to the screen when “me” is flashed to the right hemisphere—then Miller believes there must be a disconnect between the self-concept contained in each side of the brain.
Miller’s research is ongoing. But, he said, if the study finds that each hemisphere evaluates itself differently from the other, it could add a new layer of understanding to how divided the mind really is.
“Split-brain patients give you a unique glimpse into a state of consciousness you wouldn’t see otherwise,” Miller told me.
“There is something quite unique in interacting with a split-brain patient,” he added. “All the interactions you are engaging in are with left hemisphere, and you can suddenly manipulate things to interact with right hemisphere and it’s a completely different experience. A completely different consciousness.”

An official website of the United States government
The .gov means it’s official. Federal government websites often end in .gov or .mil. Before sharing sensitive information, make sure you’re on a federal government site.
The site is secure. The https:// ensures that you are connecting to the official website and that any information you provide is encrypted and transmitted securely.
- Publications
- Account settings
Preview improvements coming to the PMC website in October 2024. Learn More or Try it out now .
- Advanced Search
- Journal List
- Front Hum Neurosci
- PMC10667445
Visual integration across fixation: automatic processes are split but conscious processes remain unified in the split-brain
1 Department of Psychology, University of Amsterdam, Amsterdam, Netherlands
2 Amsterdam Brain and Cognition (ABC) Center, University of Amsterdam, Amsterdam, Netherlands
Maria-Chiara Villa
3 Department of Psychology, University of Turin, Turin, Italy
Sabrina Siliquini
4 Child Neuropsychiatry Unit, Marche Polytechnic University, Ancona, Italy
Gabriele Polonara
5 Department of Odontostomatologic and Specialized Clinical Sciences, Marche Polytechnic University, Ancona, Italy
Claudia Passamonti
6 Psychology Unit, Ospedali Riuniti, Ancona, Italy
Simona Lattanzi
7 Department of Experimental and Clinical Medicine, Marche Polytechnic University, Ancona, Italy
Nicoletta Foschi
8 Epilepsy Center-Neurological Clinic, Azienda “Ospedali Riuniti”, Ancona, Italy
Edward H. F. de Haan
9 Donders Institute, Radboud University, Nijmegen, Netherlands
10 St. Hugh’s College, Oxford University, Oxford, United Kingdom
Associated Data
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
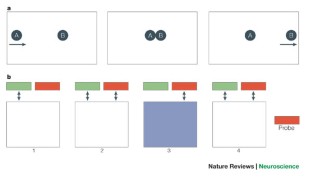
The classic view holds that when “split-brain” patients are presented with an object in the right visual field, they will correctly identify it verbally and with the right hand. However, when the object is presented in the left visual field, the patient verbally states that he saw nothing but nevertheless identifies it accurately with the left hand. This interaction suggests that perception, recognition and responding are separated in the two isolated hemispheres. However, there is now accumulating evidence that this interaction is not absolute; for instance, split-brain patients are able to detect and localise stimuli anywhere in the visual field verbally and with either hand. In this study we set out to explore this cross-hemifield interaction in more detail with the split-brain patient DDC and carried out two experiments. The aim of these experiments is to unveil the unity of deliberate and automatic processing in the context of visual integration across hemispheres. Experiment 1 suggests that automatic processing is split in this context. In contrast, when the patient is forced to adopt a conscious, deliberate, approach, processing seemed to be unified across visual fields (and thus across hemispheres). First, we looked at the confidence that DDC has in his responses. The experiment involved a simultaneous “same” versus “different” matching task with two shapes presented either within one hemifield or across fixation. The results showed that we replicated the observation that split brain patients cannot match across fixation, but more interesting, that DDC was very confident in the across-fixation condition while performing at chance-level. On the basis of this result, we hypothesised a two-route explanation. In healthy subjects, the visual information from the two hemifields is integrated in an automatic, unconscious fashion via the intact splenium, and this route has been severed in DDC. However, we know from previous experiments that some transfer of information remains possible. We proposed that this second route (perhaps less visual; more symbolic) may become apparent when he is forced to use a deliberate, consciously controlled approach. In an experiment where he is informed, by a second stimulus presented in one hemifield, what to do with the first stimulus that was presented in the same or the opposite hemifield, we showed that there was indeed interhemispheric transfer of information. We suggest that this two-route model may help in clarifying some of the controversial issues in split-brain research.
- Two experiments with the split-brain patient DDC.
- Experiment I shows that DDC is very poor at matching two stimuli presented across fixation but he feels nevertheless very confident about his performance.
- Experiment II shows that DDC can combine information presented separately in his two hemifields when he is forced to carry out a task in a deliberate, sequential manner.
- We propose a two-route model for the interhemispheric transfer of visual information. The first route is fast, automatic and unconscious integration of the two hemifields via the splenium. The second is a slow deliberate, conscious integration process.
Introduction
The corpus callosum is the main connection between the two hemispheres (e.g., Innocenti, 1986 ; Gazzaniga, 2000 ; Wahl et al., 2007 ). In “split-brain” patients, the corpus callosum has been surgically cut to alleviate severe epilepsy that does not respond to medication. This operation may cause a curious phenomenon. When a picture of an object is presented in the right visual hemifield, the patient responds correctly with the right hand and verbally. However, when the object is presented in the left hemifield the patient verbally states that he/she saw nothing but nevertheless identifies the object accurately with the left hand only, for instance, by making a drawing of the object ( Gazzaniga et al., 1962 ; Gazzaniga, 1967 , 1998 ; Sperry, 1968 , 1984 ; Wolman, 2012 ). This is in agreement with the human anatomy; the right hemisphere receives visual input from the left hemifield and controls the left hand, and vice versa ( Penfield and Boldrey, 1937 ; Cowey, 1979 ; Sakata and Taira, 1994 ). Moreover, the left hemisphere is generally the site of language processing ( Ojemann et al., 1989 ; Vigneau et al., 2006 ). Thus, it seems that resection of the corpus callosum causes each hemisphere to gain its own conscious awareness ( Sperry, 1984 ). The left hemisphere is only aware of the right hemifield and can demonstrate this through its control of the right hand and verbal capacities, while the right hemisphere is only aware of the left hemifield, which it expresses through its control of the left hand.
On closer examination, this classic ‘response x visual field’ interaction appears less than absolute. First, Sperry (1968) himself already concluded that there are clear exceptions. Second, there are a number of earlier studies that failed to observe this interaction and found that responding was well-above chance with both hands (e.g., Levy et al., 1972 ; Kingston, 1994 ; Corballis, 1995 ). More recently, we ( Pinto et al., 2017a ) performed a quantitative study into this interaction. Rather than relying on qualitative summaries, we employed a quantitative approach. For this goal we had a substantial number of trials in each condition, forced-choice responding, and a large number of different stimuli. Moreover, we employed advanced fixation control with an eye-tracker,. The response type (left hand, right hand or verbally) was varied systematically. We found, in two split-brain patients, that although visual field played a major role in most tasks, a ‘response type x visual field’ interaction was never observed. This result held across all tasks (detection, localization, orientation matching, labelling and visual matching), and all tested types of stimuli ([isoluminant] dots, simple shapes, oriented rectangles, pictures of objects). Performance was always well-above chance and comparable with respect to accuracy across the three response modalities. Similar observations were recently reported by de Haan et al. (2020a) for the detection and localisation of tactile stimulation. Nevertheless, the information transfer between the two hemispheres in split-brain patients remains controversial. For instance, Volz and Gazzaniga (2017) have suggested that these effects might be caused by confounds such as ipsilateral arm control and/or cross-cueing. Ipsilateral arm control can be disregarded as an alternative explanation. There is very little evidence supporting the suggestion that ipsilateral control can support fine-grained distal movements of the arm required for pointing. The latter suggestion, i.e., cross-cueing, refers to the possibility that one hemisphere can inform the other hemisphere via subtle cues, such as touching the other hand or even via movements of the tongue in the mouth. In response, we have argued that this is an unlikely explanation ( Pinto et al., 2017b ). The main point being the fact that split-brain patients have no reason to develop intricate cross-cueing strategies as they do not experience problems in everyday life; with naturally occurring head- and eye-movements both hemispheres are fully informed about the outside visual world. In addition, such cueing is very limited in information load, probably not more than 1 bit, and most tasks that we used ( Pinto et al., 2017a ) required a more extensive information transfer.
Another observation in split-brain patients, that is not contested, concerns the fact that they are very poor at matching stimuli that are presented across the point of fixation. For instance, if two shapes or pictures of objects are presented for a “same” versus “different” decision with one in each hemifield, patients perform at chance level. Taking these two strands of evidence together, Pinto et al. (2017c) suggested subsequently that, although matching across fixation is no longer possible and the corpus callosum is thus necessary for integration of visual information from the two hemifields, split-brain patients continue to function as having a single mind and are able to respond using all response modalities.
Obviously, there is a paradox here that requires further investigation. If the single mind has access to information from both hemispheres (or to put it differently while remaining closer to the data; if both hemispheres – each controlling one hand – have access to information from both hemispheres), how come that a split-brain patient cannot match stimuli across fixation? One possible factor that might be helpful here is the distinction between automatic processing that does not necessarily provokes, or relies on, a conscious experience versus conscious, deliberate processing. There is abundant evidence for the position that to a substantial extent cortical processing of visual information proceeds without conscious awareness (e.g., de Gelder et al., 2001 ; de Haan et al., 2021 ). Therefore, the first question that we address here is to what extent is a split-brain patient consciously aware of the visual information in a simple matching task. In order to so, we developed a simultaneous matching task with both stimuli presented within one hemifield (left or right) or across fixation with one stimulus in each hemifield and using confidence ratings as a proxy measure for conscious awareness.
Case description
Patient DDC is a classic “split-brain” patient who also participated in recent studies by Pinto et al. (2017a , 2020) and de Haan et al. (2020b , 2021) . In order to treat his medication resistant epilepsy, his corpus callosum was completely removed and most of the anterior commissure. Note that other than the resection of the corpus callosum, DDC has no brain damage, and he falls within the normal IQ range. See Corballis et al. (2010) , Pinto et al. (2020) , and Pizzini et al. (2010) for detailed clinical and radiological descriptions of this patient.
Tracking eye movements
In both experiments, we measured eye movements with an Eyelink 1,000 (SR research, Mississauga, Ontario, Canada). In Experiment 1, trials were excluded from further analysis if a saccade – with a horizontal amplitude larger than 40 pixels (40 pixels <1.31° of visual angle) – occurred, after the experimenter started the trial and the stimuli had disappeared, or if there were any missing eye positions in this interval. In Experiment 2, trials were excluded if a saccade – with a horizontal amplitude larger than 40 pixels (40 pixels <1.31° of visual angle) – occurred, after the experimenter started the trial and the first stimuli had disappeared, or after the experimenter continued the trial and the second stimuli had disappeared with a horizontal amplitude, or if there were any missing eye positions during these intervals.
This is a conservative measure of saccades (generally eye movements up to 2° of visual angle are considered micro-saccades). Finally, in both experiments, at the start of the trial, the absolute measured horizontal eye position had to be within 80 pixels of the objective horizontal center of the screen. Again a conservative measure, since the only item on the screen at the start of the trial is the fixation spot. It is quite unlikely, even difficult to fixate anywhere else than at this fixation spot.
In both Experiment 1 and Experiment 2, we employed permutation testing for statistical testing, and a significance level of 0.05.
Experiment I: Same-different matching
Materials and stimuli.
The set-up of Experiment 1 was as follows (see Figure 1 ). First, the patient fixated the fixation spot at the center of the screen (diameter: 0.44°, CIE x,y coordinates: 0.284, 0.311, luminance: 20.8 cd/m 2 ) against a dark gray background (CIE x,y coordinates: 0.279, 0.304, luminance: 5.56 cd/m 2 ). Subsequently, two shapes appeared. These shapes were identical on 50% of the trials, and different on the other 50%. The possible shapes were a black square (width and height: 1.11°), or a black circle (diameter: 1.28°) or a black triangle (width: 2.1°, height: 1.81°) appeared either 21° to the left or 7° to the right of the center of the fixation spot. All shapes were equally likely to appear. The two stimuli either appeared both in the left visual field (one stimulus 21° to the left of fixation, the other stimulus 7° to the left of fixation), around fixation (one stimulus 7° to the left of fixation, the other stimulus 7° to the right of fixation), or both in the right visual field (one stimulus 7° to the right of fixation, the other stimulus 21° to the right of fixation). This display was presented for 0.12 s. After this display a response screen was presented in which the patient indicated whether the two shapes were the same or different. This was followed by a confidence judgment where the patient indicated his confidence (from 1 to 3 where 1 indicated low confidence and 3 high confidence).

In Experiment 1 two stimuli appeared. Both stimuli either appeared in the left visual field, the right visual field, or around fixation (as depicted here). The participant first indicated whether both stimuli were the same. Subsequently he indicated his confidence in his judgment on a 3-point scale, where 1 indicated guess and 3 indicated certainty.
Stimuli were presented on a 24-in. monitor set to a resolution of 1920 × 1,080 at a refresh rate of 60 Hz controlled by a Dell Optiplex 760 computer (Dell, Dallas, TX) running Windows 8. The experiment was programmed in Matlab 7.7.0 (The Mathworks Company, Natick, MA) using the Psychophysics Toolbox routines ( Brainard and Vision, 1997 ; Pelli, 1997 ). The patient was seated approximately 50 centimeters away from the screen. He rested his chin on a headrest to minimize head movements.
The trial was started manually by the experimenter. The experimenter, who could not see the stimulus screen, employed the eyelink to ensure that DDC fixated the center of the screen. Only if this was the case, the experimenter started the trial by pressing the space bar. Two stimuli appeared either within one hemifield, or across both hemifields. First DDC indicated whether both stimuli were the same or different. Subsequently he indicated his confidence in his judgment from 1 (guess) to 3 (certain). On half of the trials both stimuli appeared in one hemifield (equally often in the left and the right visual hemifield) and on the other half of the trials one stimulus appeared in each hemifield. Furthermore, on half of the trials both stimuli were the same, and on the other half of the trials both stimuli were different. All conditions were randomly intermixed. The experiment consisted of 3 blocks of 48 trials. DDC responded verbally (“same” vs. “different”) throughout the entire experiment.
See Figures 2 , ,3 3 for an overview of the results of Experiment 1. If both stimuli appeared within one hemifield, DDC’s performance exceeded chance performance ( p = 0.001), however when both stimuli appeared across fields, performance was not better than chance ( p = 0.5). When both stimuli appeared in one hemifield, performance was better than chance, both on low confidence trials ( p = 0.03) and high confidence trials (p = 0.001). However, when stimuli appeared across hemifields performance was not better than chance, both on low confidence and high confidence trials (ps > 0.43). Performance on high confidence trials was more accurate than on low confidence trials if both stimuli appeared within one hemifield ( p < 0.05), but not when both stimuli appeared across hemifields ( p = 0.73).

An overview of the results of Experiment 1. DDC was more accurate when the stimuli within one hemifield than when the stimuli appeared across hemifields. In the latter case, performance did not exceed random guessing. Moreover, only if both stimuli appeared within one hemifield, DDC was more accurate when he was more confident.

Incidence of low and high confidence trials depending on whether both stimuli appeared with or across hemifields. Remarkably, DDC was generally more confident when both stimuli appeared in different hemifields.
The main finding here is that despite worse (in fact, chance) performance when both stimuli appeared across hemifields rather than within one hemifield, the incidence of high confidence trials was higher in the former case. If both stimuli appeared across hemifields, DDC was highly confident on 52% of the trials. Yet, if both stimuli appeared within one hemifield DDC was highly confident on 21% of the trials. This difference was significant ( p < 0.001). Note that in healthy adults the across hemifields condition is generally experienced as easier, since both stimuli appear more centrally in the visual field than if the stimuli appear within one hemifield. Remarkably, DDC had the same subjective impression – if stimuli appear centrally, the task ‘felt’ easier – despite the objective breakdown of his performance. In other words, DDC seemed to be completely unaware of his poor performance when the stimuli appeared in different visual fields. Anecdotally, during the experiment DDC remarked that he found the experiment much easier if the stimuli appeared around fixation (i.e., across visual hemifields).
First, this experiment replicated earlier studies (e.g., Pinto et al., 2017a ) that had demonstrated that within-hemifield comparisons can be carried out accurately but that matching across the point of fixation is at chance-level. In addition, the accuracy of his within-hemifield matching is highly correlated with his confidence score, suggesting that he is consciously perceiving both stimuli and aware of the matching process. Thus, we conclude that he is not reacting in a reflexive fashion. However, and this is the most intriguing result, when the two stimuli were presented across hemifields, he was highly confident despite the fact his performance was at chance-level. In fact, he was more confident in the across- than in the within-hemifields conditions. The absence of a correlation between accuracy and confidence in the across-hemifields condition suggests that DDC is completely oblivious of his inability to compare stimuli that have been presented in his two hemifields.
There are, at least, two possible explanations for this observation. The first explanation is that the resection of the corpus callosum has resulted in an additional deficit affecting the introspective insight into his own functioning. In other words, it is a specific form of anosognosia. Although we cannot rule out this explanation, there is a second, perhaps more likely, explanation that concerns the possibility that automatic, unconscious visual integration across fixation is split but conscious processes remain unified in a split-brain patient. This hypothesis is based on the idea that the information from the two hemifields is integrated, via the corpus callosum, in an automatic, unconscious fashion in healthy observers. This fits the normal introspection; we do not experience a vertical line in the middle of our field of vision. It also fits with classic observation that split-brain patients generally feel unchanged (c.f. Bogen et al., 1965 ). Of course, in everyday life, they do not have a problem because they can make eye and head movement. As a result of his conviction that he perceives an integrated and complete visual field of vision, DDC feels confident in his visual matching but he is let down by the broken automatic integration of visual information across fixation.
If this explanation is correct, then it follows that the observation that a split-brain patient is able to respond accurately and confidently with both hands and verbally to stimuli presented anywhere in the visual field is based on a different, unified and perhaps conscious process. This process may be characterised as less visual, more symbolic in nature and may be communicated via subcortical pathways. In order to test this hypothesis, we developed the following experiment. It is aimed at the question whether he can combine visual information from the two hemifields in a conscious and deliberate fashion. Therefore, we adapted the first experiment in such a way that he now must process the two shapes separately.
Experiment II: Integration across hemifields
The set-up of Experiment 2 was as follows (see Figure 4 ). First, the patient fixated the fixation spot diameter: 0.58° (CIE x,y coordinates: 0.284, 0.311, luminance: 20.8 cd/m 2 ) at the center of the screen against a gray background (CIE x,y coordinates: 0.282, 0.309, luminance: 13 cd/m 2 ). Subsequently, a black square (width and height: 1.11°) or a black circle (diameter: 1.28°) appeared either 13.4° to the left or 13.4° to the right of the center of the fixation spot. This display was presented for 0.12 s. Subsequently, only the fixation spot was presented until the experimenter pressed a spacebar. The experimenter did so when he ensured that the participant was fixating the center of the screen. The experimenter monitored the eye movements of the participant during the experiment; however, he could not see the stimulus screen. Finally, either a question mark (width and height: 1.11°, CIE x,y coordinates: 0.405, 0.521, luminance: 54.5 cd/m 2 ) or an “X” (width and height: 1.11°, CIE x,y coordinates: 0.641, 0.341, luminance: 11.5 cd/m 2 ) was presented either 13.4° to the left or 13.4° to the right of the center of the fixation spot. The final display was presented for 0.12 s. This was followed by the response screen where DDC could select one of three options – a square, a circle, or an “X”.

Initially either a square or a circle appeared in the left or the right visual field. Subsequently either a question mark or an “X” appeared, again either in the left or the right visual field. If the second stimulus was an “X” then the participant had to select an “X” as response. If the second stimulus was a question mark, then the participant had to select the first stimulus he saw. Thus, in this example, the square is the correct response.
The trial was started manually by the experimenter. The experimenter, who could not see the stimulus screen, employed the eyelink to ensure that DDC fixated the center of the screen. Only if this was the case, the experimenter started the trial by pressing the space bar. Subsequently either a circle or square appeared for 0.12 s, either to the left or right of fixation, followed by a fixation spot at the center of the screen. At this point the experimenter again employed the eyelink to ensure that DDC fixated the center of the screen. Again, only if this was the case the experimenter continued the trial by pressing the space bar. Then either a question mark or an “X” appeared for 0.12 s, either to the left or right of fixation. Finally, the response display, consisting of a square, a circle and an “X” appeared and remained visible until DDC responded. The task of DDC was as follows. If the second stimulus was an “X” then he had to select an “X” from the response screen. However, if the second stimulus was a question mark, then he had to indicate what he had seen in the first display, by either selecting a square or a circle.
The experiment consisted of 4 blocks of 48 trials. There were four conditions, the shape either appeared in the left or the right visual field, followed by a second stimulus which again appeared either in the left or the right visual field – the second stimulus was either an “X” or a question mark. All conditions occurred equally often and were randomly intermixed throughout the experiment. The shape was equally likely to be a square or a circle, the second stimulus was equally likely to be an “X” or a question mark. Throughout the experiment DDC responded verbally.
In a control experiment we verified that DDC’s performance on directly comparing stimuli across the midline (same stimuli and same positions of the stimuli as in Experiment 2) did not exceed chance performance. This was true for all four stimuli. Thus, for example, DDC was no better than chance in indicating if the stimuli were the same or different if either two question marks or two squares appeared in LVF and RVF, or a question mark appeared in one visual field, and a square appeared in the other visual field. Moreover, DDC was highly accurate when he compared these stimuli to each other when they both appeared in one visual hemifield.
The main result of Experiment 2 is that when the first stimulus and the second stimulus appeared in different fields, so that DDC had to make a deliberate delayed comparison across hemifields, he was highly accurate, see Figure 5 (Across hemifields accuracy: 80.8%, within hemifield accuracy: 74.2%. In both cases, performance was significantly better than chance, ps < 0.001. No significant difference between across and within hemifield accuracy, p = 0.19). Arguably, on the trials on which an “X” appeared as the second stimulus no across field integration was required, since DDC could simply ignore the first stimulus and select the “X” from the response set. Therefore, we performed the same analysis as before, but now we only included trials on which the second stimulus was a question mark. We again observed the same pattern as before. DDC was highly accurate even when stimuli appeared across hemifields, see Figure 5 (Across hemifields accuracy: 68.7%, within hemifield accuracy: 58.7%. In both cases performance was significantly better than chance, ps < 0.001. No significant difference was apparent between across and within hemifield accuracy, p = 0.29).

An overview of the results of Experiment 2. In this experiment DDC had to deliberately integrate information. Now his performance was at least as good in the across hemifields as in the within hemifield condition. Moreover, in all conditions his performance was well above chance.
Crucially, in this experiment DDC’s was performing very well in all conditions. This was especially noteworthy when the second stimulus was a question mark, and the first and the second stimulus appeared in opposite hemifields. The main difference with experiment I is that he is now forced to pay active attention to the first stimulus: he might later on be asked whether it was a “□” or a “○”. This deliberate processing results in an almost perfect recollection of the first stimulus irrespective of whether this stimulus was presented in the same or the other hemifield. Thus, two hemispheres that are no longer connected via a corpus callosum interact to the degree that when one hemisphere is confronted with question mark, it can respond accurately to the question which shape had been presented to the other hemisphere.
General discussion
In this study, we focussed on an important paradox observed in split-brain patients. On the one hand, split-brain patient cannot match stimuli across fixation, while on the other hand, it has now been established that these patients can respond accurately to stimuli anywhere in the visual field with either hand or verbally. Note: this paradox stands whatever explanation one wants to entertain to explain these effects.
Our starting point for addressing this issue was to look at the distinction between automatic processing that does not necessarily provoke, or rely on, a conscious experience versus conscious, deliberate processing. In Experiment 1, we used confidence ratings as a proxy for conscious awareness. In contrast to matching stimuli presented in the same hemifield, where his performance correlated with confidence (suggesting that he was well aware of what he was doing), there was no such correlation in the condition where he was asked to match across the point of fixation. In fact, while he performs at chance-level, he is extremely confident.
We hypothesised that in the healthy brain information from the two hemifields is integrated, via the corpus callosum, in an automatic, unconscious fashion. Due to the resection of the corpus callosum, this link has been severed in a split-brain patient. However, DDC has no reason to doubt his abilities in this respect. Just as in normal introspection (we experience an integrated field of vision), DDC also subjectively perceives an integrated and complete visual world under normal circumstances when he can move his head eyes. However, this automatic integration of visual information itself is blocked by the resection of the splenium. It is not immediately clear on the basis of what information he comes to the (often incorrect) response. Perhaps this is the result of a completion process across fixation (e.g., Sergent, 1988 ; McCarthy et al., 2006 ).
If this explanation is correct, then it follows that the observation that a split-brain patient is able to respond accurately and confidently with both hands and verbally to stimuli presented anywhere in the visual field is based on a different, unified and perhaps conscious process. We speculated that this process is characterised by less visual detail and more symbolic in nature, and that it might involve subcortical pathways. In order to test this hypothesis, we adapted the first experiment in such a way that he now has to process the two shapes separately. The results showed convincingly that under these experimental conditions, DDC was as accurate in combining information presented in two different hemifields as he as when the information was presented in the same hemifield.
It could be argued that the results of Experiment 2, where we aimed to force the split-brain patient to consciously integrate information across hemifields, were not caused by conscious effort, but by the delay between the first stimulus and the second stimulus. In other words, maybe the effect was due to changing the task from an immediate to a delayed matching task. Perhaps this temporal gap allowed for more cross-cueing for instance. However, note that such an explanation is unlikely to be true, as previous research has shown that a (partially) split-brain patient was not able to perform a delayed matching task either ( Funnell et al., 2000 ). In other words, it seems that if a split-brain patient relies on automatic integration of information across visual fields, performance plummets, even if a temporal gap is added. The patient is only successful if he consciously integrates the visual information coming from both hemifields.
So, we propose a two-route explanation for these observations. In healthy subjects, the visual information from the two hemifields is integrated in an automatic, unconscious fashion via the intact splenium. It is fast and integrates detailed visual information but there is no possibility for introspection or control. This route has been blocked in a split-brain patient. However, the transfer of information remains possible when the patient is forced to use a deliberate, consciously controlled approach. The observation that he does not use this approach spontaneously is due to his false belief that he can rely on the first route in matching tasks. There are, at least, two questions that follow from this proposal. First, what is the nature of the information that is transferred via the second route? We suggest that it is more abstract, for instance the concept of “circle” or a “square”, or even the verbal label. As we used a response screen showing the different options and he responded verbally, we cannot distinguish between these possible options. The second question concerns the neural basis for this spared interhemispheric transfer. Several authors ( Pinto et al., 2017c ; Corballis et al., 2018 ; de Haan et al., 2020b ) suggested that these effects could be the result of intact subcortical pathways. This is supported by the observation by Savazzi et al. (2007) , who showed that the superior colliculus is likely to play a role in the visual transfer between the hemispheres. However, at present, it is not possible to exclude alternative explanations, such as quantum processing (e.g., Chalmers and McQueen, 2020 ) or complex cross-cueing. The latter is not very likely, as in the Experiment II it would have to be the responding hemisphere that has to probe the other hemisphere in half the trails (and not in the half). Such a deliberate and controlled strategy would be obvious to the patient. In our experience, patients have not commented on using such strategies.
In two experiments, we found that automatic integration of visual information does not occur in the split-brain, yet deliberate integration remains possible. Moreover, the split-brain patient was unaware of this failure of automatic integration. Together these findings support the view that in a split-brain the conscious mind remains unified while automatic, unconscious processing is divided. We suggest that our proposal for a two-route model may help in clarifying some of the controversial issues in split-rain research.
Data availability statement
Ethics statement.
Ethical approval was not required for the studies involving humans because This was part of standard research with the subjects. The studies were conducted in accordance with the local legislation and institutional requirements. Written informed consent for participation was not required from the participants or the participants’ legal guardians/next of kin in accordance with the national legislation and institutional requirements because this was part of standard research with the subjects.
Author contributions
YP: Conceptualization, Methodology, Supervision, Writing – original draft, Writing – review & editing. M-CV: Writing – original draft, Writing – review & editing. SS: Writing – review & editing. GP: Writing – review & editing. CP: Writing – review & editing. SL: Writing – review & editing. NF: Writing – review & editing. EH: Methodology, Writing – original draft, Writing – review & editing.
Acknowledgments
We would like to thank Gabriella Venanzi and Pamela Rosettani for their cooperation in this research.
Funding Statement
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by a European Research Council Advanced grant FAB4V (#339374) to EH, and a Templeton grant (ID #61382, “Towards understanding a unified mind”) to YP.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
- Bogen J. E., Fisher E., Vogel P. (1965). Cerebral commissurotomy: a second case report . J. Am. Med. Assoc. 194 , 1328–1329. [ PubMed ] [ Google Scholar ]
- Brainard D. H., Vision S. (1997). The toolbox of psychophysics . Spatial vision 10 , 433–436. [ PubMed ] [ Google Scholar ]
- Chalmers D. J., McQueen K. J. (2020). “ Consciousness and the collapse of the wave function ” in Consciousness and quantum mechanics . ed. Gao S. (Oxford University Press; ). [ Google Scholar ]
- Corballis M. C. (1995). Visual integration in the split brain . Neuropsychologia 33 , 937–959. doi: 10.1016/0028-3932(95)00032-X, PMID: [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Corballis M. C., Birse K., Paggi A., Manzoni T., Pierpaoli C., Fabri M. (2010). Mirror-image discrimination and reversal in the disconnected hemispheres . Neuropsychologia 48 , 1664–1669. doi: 10.1016/j.neuropsychologia.2010.02.011, PMID: [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Corballis M. C., Corballis P. M., Berlucchi G., Marzi C. A. (2018). Perceptual unity in the split brain: the role of subcortical connections . Brain 141 :e46. doi: 10.1093/brain/awy085 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Cowey A. (1979). Cortical maps and visual perception the grindley memorial lecture . Q. J. Exp. Psychol. 31 , 1–17. doi: 10.1080/14640747908400703, PMID: [ PubMed ] [ CrossRef ] [ Google Scholar ]
- de Gelder B., Heywood B. C. A., De Haan E. H. F. (2001). Varieties of unconscious processing . Oxford University Press. [ Google Scholar ]
- de Haan E. H. F., Corballis P. M., Hillyard S. A., Marzi C. A., Seth A., Lamme V. A. F., et al.. (2020b). Split-brain: what we know now and why this is important for understanding consciousness . Neuropsychol. Rev. 30 , 224–233. doi: 10.1007/s11065-020-09439-3, PMID: [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- de Haan E. H. F., Fabri M., Dijkerman H. C., Foschi N., Lattanzi S., Pinto Y. (2020a). Unified tactile detection and localisation in split-brain patients . Cortex 124 , 217–223. doi: 10.1016/j.cortex.2019.11.010 [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- de Haan E. H. F., Scholte H. S., Pinto Y., Foschi N., Polonara G., Fabri M. (2021). Singularity and consciousness: a neuropsychological contribution . J. Neuropsychol. 15 , 1–19. doi: 10.1111/jnp.12234, PMID: [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Funnell M. G., Corballis P. M., Gazzaniga M. S. (2000). Insights into the functional specificity of the human corpus callosum . Brain 123 , 920–926. doi: 10.1093/brain/123.5.920, PMID: [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Gazzaniga M. S. (1967). The split brain in man . Sci. Am. 217 , 24–29. doi: 10.1038/scientificamerican0867-24, PMID: [ CrossRef ] [ Google Scholar ]
- Gazzaniga M. S. (1998). “ The problem of consciousness ” in Cognitive neuroscience – the biology of the mind W . eds. Gazzaniga M. S., Ivry R. B., Mangun G. R. (New York, NY, USA: W. Norton & Company; ), 542–543. [ Google Scholar ]
- Gazzaniga M. S. (2000). Cerebral specialization and interhemispheric communication: does the corpus callosum enable the human condition? Brain 123 , 1293–1326. doi: 10.1093/brain/123.7.1293, PMID: [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Gazzaniga M. S., Bogen M. S., Sperry R. W. (1962). Some functional effects of sectioning the cerebral commissures in man . Proc. Natl. Acad. Sci. U. S. A. 48 , 1765–1769. doi: 10.1073/pnas.48.10.1765, PMID: [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Innocenti G. M. (1986). “ General organization of callosal connections in the cerebral cortex ” in Sensory-motor areas and aspects of cortical connectivity . eds. Jones E.G., et al.. (New York: Plenum press, Springer; ), 291–353. [ Google Scholar ]
- Kingston A. (1994). Covert orienting in the split brain reveals hemispheric specialization for object-based attention . Psychol. Sci. 5 , 380–383. [ PubMed ] [ Google Scholar ]
- Levy J., Trevarthen C., Sperry R. W. (1972). Perception of bilateral chimeric figures following hemispheric deconnexion . Brain 95 , 61–78. doi: 10.1093/brain/95.1.61, PMID: [ PubMed ] [ CrossRef ] [ Google Scholar ]
- McCarthy R. A., James-Galton M., Plant G. T. (2006). Form completion across a hemianopic boundary: Behindsight? Neuropsychologia 44 , 1269–1281. doi: 10.1016/j.neuropsychologia.2006.01.036, PMID: [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Ojemann G., Ojemann J., Lettich E., Berger M. (1989). Cortical language localization in left, dominant hemisphere: an electrical stimulation mapping investigation in 117 patients . J. Neurosurg. 71 , 316–326. doi: 10.3171/jns.1989.71.3.0316, PMID: [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Pelli D. G. (1997). The VideoToolbox software for visual psychophysics: transforming numbers into movies . Spat. Vis. 10 , 437–442. doi: 10.1163/156856897X00366, PMID: [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Penfield W., Boldrey E. (1937). Somatic motor and sensory representation in the cerebral cortex of man as studied by electrical stimulation . Brain 60 , 389–443. doi: 10.1093/brain/60.4.389 [ CrossRef ] [ Google Scholar ]
- Pinto Y., de Haan E. H. F., Lamme V. A. F. (2017c). The Split-brain phenomenon revisited: a single conscious agent with Split perception . Trends Cogn. Sci. 21 , 835–851. doi: 10.1016/j.tics.2017.09.003 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Pinto Y., de Haan E. H. F., Villa M.-C., Siliquini S., Polonara G., Passamonti C., et al.. (2020). Unified visual working memory without the anterior Corpus callosum . Symmetry 12 :2106. doi: 10.3390/sym12122106 [ CrossRef ] [ Google Scholar ]
- Pinto Y., Lamme V. A., de Haan E. H. (2017b). Cross-cueing cannot explain unified control in split-brain patients . Brain 140 :e68. doi: 10.1093/brain/awx235 [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Pinto Y., Neville D. A., Otten M., Corballis P. M., Lamme V. A., de Haan E. H. F., et al.. (2017a). Split brain: divided perception but undivided consciousness . Brain 140 , 1231–1237. doi: 10.1093/brain/aww358, PMID: [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Pizzini F. B., Polonara G., Mascioli G., Beltramello A., Foroni R., Paggi A., et al.. (2010). Diffusion tensor tracking of callosal fibers several years after callosotomy . Brain Res. 1312 , 10–17. doi: 10.1016/j.brainres.2009.11.030, PMID: [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Sakata H., Taira M. (1994). Parietal control of hand action . Curr. Opin. Neurobiol. 4 , 847–856. doi: 10.1016/0959-4388(94)90133-3, PMID: [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Savazzi S., Fabri M., Rubboli G., Paggi A., Tassinari C. A., Marzi C. A. (2007). Interhemispheric transfer following callosotomy in humans role of the superior colliculus . Neuropsychologia 45 , 2417–2427. doi: 10.1016/j.neuropsychologia.2007.04.002, PMID: [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Sergent J. (1988). An investigation into perceptual completion in blind areas of the visual field . Brain 111 , 347–373. doi: 10.1093/brain/111.2.347. PMID: 3378140, PMID: [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Sperry R. W. (1968). Hemisphere deconnection and unity in conscious awareness . Am. Psychol. 23 :723. doi: 10.1037/h0026839, PMID: [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Sperry R. (1984). Consciousness, personal identity and the divided brain . Neuropsychologia 22 , 661–673. doi: 10.1016/0028-3932(84)90093-9, PMID: [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Vigneau M., Beaucousin V., Hervé P. Y., Duffau H., Crivello F., Houdé O., et al.. (2006). Meta-analyzing left hemisphere language areas: phonology, semantics, and sentence processing . NeuroImage 30 , 1414–1432. doi: 10.1016/j.neuroimage.2005.11.002, PMID: [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Volz L. J., Gazzaniga M. S. (2017). Interaction in isolation: 50 years of insights from split-brain research . Brain 140 , 2051–2060. doi: 10.1093/brain/awx139, PMID: [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Wahl M., Lauterbach-Soon B., Hattingen E., Jung P., Singer O., Volz S., et al.. (2007). Human motor corpus callosum: topography, somatotopy, and link between microstructure and function . J. Neurosci. 27 , 12132–12138. doi: 10.1523/JNEUROSCI.2320-07.2007, PMID: [ PMC free article ] [ PubMed ] [ CrossRef ] [ Google Scholar ]
- Wolman D. (2012). A tale of two halves . Nature 483 , 260–263. doi: 10.1038/483260a, PMID: [ PubMed ] [ CrossRef ] [ Google Scholar ]
A Wealth of Free Psychology!
Sperry (1968) – split brain study.
Sperry, R. W. (1968). Hemisphere deconnection and unity in conscious awareness. American Psychologist , 23 (10), 723.
This is the classic biological psychology study which you will look at for your H167 AS OCR Psychology exam. You will also need this study for your OCR H567 A Level Psychology core studies exam.
The theme of the biological psychology studies in the H167 exam is regions of the brain . This study by Sperry (1968) has the tagline by OCR: Split-brain Study .
What does it mean to have a ‘split-brain’?
The brain is composed of two cerebral hemispheres: the left hemisphere and the right hemisphere. These hemispheres are connected in the brain by the corpus callosum , and other smaller connections, but we need not worry about them here. Having a ‘split-brain’ simply means that the corpus callosum has been severed.
What is the corpus callosum?
The corpus callosum is a large band of white matter (white matter is efficient at sending information), which connects the two hemispheres of the brain.
Why would the corpus callosum be severed?
The corpus callosum may be severed by surgeons in order to reduce the symptoms of epilepsy. In epilepsy, on hemisphere of the brain is usually responsible. Very simply put, when an epileptic episode occurs, there is an electrical storm in one hemisphere of the brain, which then travels across the corpus callosum, causing the entire brain to be affected and then a blackout occurs. By severing the corpus callosum this travelling of the electrical storm cannot occur and thus blackout and epileptic fits cease.
The severing of the corpus callosum is called a commissurotomy .
Difficulties learning Sperry’s Study
One of the most confusing and challenging parts for most students when learning Sperry’s study is the idea of contralateralisation.
The brain is composed of two hemispheres, as mentioned earlier. Most stimuli (sound is more complex) is processed contralaterally. This means that if stimuli enters on the left, for example, the left hand, it is processed in the right hemisphere and visa versa.
For the most part, language is processed in the left hemisphere. (For the most part because this gets complicated, especially when participants’ have different dominant hands, but for the purposes of this study, remember that language, especially spoken language is processed in the left side of the brain).
These two ideas are quite difficult to remember, as it quickly becomes confusing. I suggest testing each other in small groups in order to get to grips with these ideas.
If you want to read more on the work of Roger Sperry, then read: Beyond a World Divided: Human Values in the Brain-mind Science of Roger Sperry
Aim of the Experiment
Method and design.
Sperry (1968) used a quasi experiment in a laboratory with an independent measures design.
The independent variable was whether the individual had a split-brain or not.
There were dependent variable was that individual’s performance on visual and tactile tasks.
Please note, Sperry’s (1968) study did not have a control group. This was because the effects of not having a split-brain were already known. Therefore, there was no need to have a control group.
Further, some have argued that this study was actually a small collection of case studies.
Sample and Sampling Method
11 Participants.
All the participants were epileptics had previously undergone commissurotomies to deal with their severe epileptic convulsions.
The first patient (a man) had his surgery over 5½ years before the study was conducted.
The second patient, a housewife and mother in her 30s had her surgery more than 4 years before the study was conducted.
The other 9 patients had their surgery at varying times but not long before the study was conducted.
Visual fields.
We have two visual fields. These visual fields should not be confused with our two eyes. We don’t need to go into the biological factors underlying the visual fields here. The left visual field, as you might guess, is what we can see on our left and the right visual field is what we can see on our right.
As you can see in the diagram, the information from the right visual field goes to the left hemisphere (see the hand with the pencil).
Visual Tasks
Sperry used a tachistoscope to present visual stimuli to the participants.
The tachistoscope has a focal point in the middle and two areas where stimuli was presented. The participants using the tachistoscope would have one eye covered and were instructed to stare at the focal point.
Information presented to the left of the focal point would be seen in the left visual field which would then travel to the right hemisphere.
Wearing an eye patch and staring at the focal point were controls. These controls ensure that stimuli was presented only to the desired visual field.
All visual stimuli was presented for only 0.1 seconds. This was another control as it is too quick for eye movements to cause visual information to enter both visual fields.
$/? Task. The $/? task had both a ‘$’ and a ‘?’ presented simultaneously (one to the right visual field and the other to the left visual field). Participants were then tested on the stimuli.
Tactile Tasks
Notice in the above image of the tachistoscope there are some objects behind the screen. In the tactile tests the participants would put their hands under the tachistoscope such that they could reach the objects. The participants’ hands were then covered. This was a control to ensure there was no visual stimuli going to either hemisphere and thus could not confound upon the results. Given that the tachistoscope was in between the participants and the objects, the participants could not see the objects. Again this was a control measure to ensure only tactile stimuli was introduced to the participants.
Participants were introduced to objects by an experimenter, who placed them in the participants’ hands.
Objects placed in the right hand of the participant are processed in the left hemisphere.
Objects placed in the left hand of the participant are processed in the right hemisphere.
Participants would only recognise stimuli if the stimuli was presented again to the same visual field. If participants were shown stimuli in the right visual field, but then shown the same stimuli to the left visual field, they would claim to have not seen it before.
Information presented to the right visual field (left hemisphere) could be described in speech and writing (with the right hand). If the same information is presented to the left visual field (right hemisphere), the participant insisted he either did not see anything or that there was only a flash of light on the left side, that is, the information could not be described in speech or writing. However the participant could point with his left hand (controlled by the right hemisphere) to a matching picture / object presented among a collection of pictures / objects.
$/? task findings. If different figures were presented simultaneously to different visual field, for example ‘$’ sign to the left visual field and ‘?’ to the right visual field, the participant could draw the ‘$’ sign with his left hand but reported that he had seen a ‘?’.
Tactile Tasks
Objects placed in the right hand (left hemisphere) could be described in speech or writing (with the right hand). If the same objects were placed in the left hand (right hemisphere) participants could only make wild guesses and often seemed unaware they were holding anything.
Objects felt by one hand were only recognised again by the same hand for example objects first sensed by the right hand could not be retrieved by the left.
When two objects were placed simultaneously in each hand and then hidden in a pile of objects, both hands selected their own object and ignored the other hand’s object.
Conclusions
People with split brains have two separate visual inner worlds, each with its own train of visual images.
Split-brain patients have a lack of cross-integration where the second hemisphere does not know what the first hemisphere has been doing.
Split-brain patients seem to have two independent streams of consciousness, each with its own memories, perceptions and impulses ie two minds in one body
Sperry (1968) Evaluation
– Quasi study – as the there was no manipulation by the experimenter, establishing cause and effect becomes more difficult than a traditional experiment.
+ Ethics – as the study used a quasi methodology, Sperry did not need to manipulate anything, which is more ethical than studies which manipulate variables.
– No control group, Sperry did not use a control group, which makes it difficult to truly establish cause and effect. However, it is important to note that in this study a control group was not needed as the results of the tasks for people without split corpus callosums were already known.
– External validity – the external validity in this study may be considered low because the stimuli was selectively delivered to one hemisphere. This does not happen in real life and thus is not representative of the everyday experience of ‘split-brain’ patients. However this selective presentation of stimuli increased the internal validity because there is an increased likelihood that Sperry measured the effects of hemisphere deconnection.
Further Reading
Beyond a World Divided: Human Values in the Brain-mind Science of Roger Sperry
Psych Yogi’s Top Ten Psychology Revision Tips for the A* Student
3 thoughts on “ Sperry (1968) – Split Brain Study ”
Helped alot for my mock exams tomoz, thanks.
Helpful to prepare the presentation of split-brain study! Thanks a lot.!
No worries – glad you found it helpful, Daisy
Comments are closed.
Live revision! Join us for our free exam revision livestreams Watch now →
Reference Library
Collections
- See what's new
- All Resources
- Student Resources
- Assessment Resources
- Teaching Resources
- CPD Courses
- Livestreams
Study notes, videos, interactive activities and more!
Psychology news, insights and enrichment
Currated collections of free resources
Browse resources by topic
- All Psychology Resources
Resource Selections
Currated lists of resources
Study Notes
Biopsychology: Evaluating Split-Brain Research
Last updated 10 Apr 2017
- Share on Facebook
- Share on Twitter
- Share by Email
Here are some key evaluation points on split-brain research.
It is assumed that the main advantage of brain lateralisation is that it increases neural processing capacity (the ability to perform multiple tasks simultaneously). Rogers et al. (2004) found that in a domestic chicken, brain lateralisation is associated with an enhanced ability to perform two tasks simultaneously (finding food and being vigilant for predators). Using only one hemisphere to engage in a task leaves the other hemisphere free to engage in other functions. This provides evidence for the advantages of brain lateralisation and demonstrates how it can enhance brain efficiency in cognitive tasks.
However, because this research was carried out on animals, it is impossible to conclude the same of humans. Unfortunately, much of the research into lateralisation is flawed because the split-brain procedure is rarely carried out now, meaning patients are difficult to come by. Such studies often include very few participants, and often the research takes an idiographic approach. Therefore, any conclusions drawn are representative only of those individuals who had a confounding physical disorder that made the procedure necessary. This is problematic as such results cannot be generalised to the wider population.
Furthermore, research has suggested that lateralisation changes with age. Szaflarki et al. (2006) found that language became more lateralised to the left hemisphere with increasing age in children and adolescents, but after the age of 25, lateralisation decreased with each decade of life. This raises questions about lateralisation, such as whether everyone has one hemisphere that is dominant over the other and whether this dominance changes with age.
Finally, it could be argued that language may not be restricted to the left hemisphere. Turk et al. (2002) discovered a patient who suffered damage to the left hemisphere but developed the capacity to speak in the right hemisphere, eventually leading to the ability to speak about the information presented to either side of the brain. This suggests that perhaps lateralisation is not fixed and that the brain can adapt following damage to certain areas.
- Biopsychology
- Split Brain Research
You might also like
The biological approach, stress: what is it good for absolutely nothing.
17th January 2017
Healthy Body, Healthy Mind
30th January 2017
Plasticity & Functional Recovery: Biopsychology Webinar Video
Topic Videos
Biopsychology: The ‘Fight or Flight’ Response Explained
Biopsychology: the ‘fight or flight’ response - evaluation, biopsychology: studying the brain - evaluation of eeg & erp, approaches in psychology: as webinar video 2016, our subjects.
- › Criminology
- › Economics
- › Geography
- › Health & Social Care
- › Psychology
- › Sociology
- › Teaching & learning resources
- › Student revision workshops
- › Online student courses
- › CPD for teachers
- › Livestreams
- › Teaching jobs
Boston House, 214 High Street, Boston Spa, West Yorkshire, LS23 6AD Tel: 01937 848885
- › Contact us
- › Terms of use
- › Privacy & cookies
© 2002-2024 Tutor2u Limited. Company Reg no: 04489574. VAT reg no 816865400.
Split Brain Research
Hemispheric lateralisation: This is the ideas that the brain’s two hemispheres are responsible for different functions; that particular functions (such as language) are the responsibility of one hemisphere but not the other- the function is lateralised.
Split-brain research: This involves individuals who have had surgical separation of their brain hemispheres, in order to relieve symptoms of epilepsy. Research can reveal to what extent other brain functions are lateralised. Sperry (1968) used a procedure where an image or word was projected to the patient’s right visual field (which would be processed by the left hemisphere) and another image was projected to the left visual field (processed by the right hemisphere). In a split-brain patient, information cannot be transmitted from one hemisphere to another, so the effects of this can be studied.
Findings from split-brain research: When a picture was shown to the right visual field, the patient could describe it easily. When the image was shown to the left visual field, the patient found it difficult to describe it, or couldn’t see anything there. This is likely due to the lack of language processing ability in the right hemisphere, which processes the left visual field. The left hemisphere was unable to receive the information due to the separation of the hemispheres.
When patients were presented with a word or object in the left visual field, then had to select that object (or a related one) from a bag using their left hand without looking, they were able to do this quite well. This is despite the fact that they couldn’t describe what the object was. They could still seemingly understand what the object was, showing that the right hemisphere is involved in understanding of objects.
When patients were presented with two words simultaneously (for example, ‘door’ ‘frame’), one in each visual field, they wrote the word in the left visual field (‘door’) with their left hand, and said the word in the right visual field (‘frame’). The left hand was much better at drawing images than the right hand, despite the fact that all of the patients tested were right-handed. This shows that the right hemisphere seems to be dominant for drawing skills.
When asked to match a face from a number of other faces, the picture presented to the left visual field (processed by the right hemisphere) was consistently selected, whilst the image presented to the right visual field (left hemisphere) was ignored. This shows that the right hemisphere seems dominant in face recognition. Images presented as composites of two faces led to the left hemisphere dominating when the patient was asked to verbally describe the face (the right-hand ‘half’ of the face was described, and the left-hand half ignored), and the right hemisphere dominated when the patient had to select a matching face from a number of others.
Evaluation:
- Sperry’s research supports the conclusion that the left hemisphere is more responsible for verbal and analytical tasks, whereas the right hemisphere is better at spatial and musical tasks. This has strengthened the understanding of how the brain works.
- Sperry’s procedure was closely controlled. Patients were given eye patches, and images were flashed up for a very brief time (fractions of a second), meaning there was no possibility of looking over and using the other visual field. This strengthens the internal validity of the studies.
- The sample used by Sperry was quite small (only 11 took part in all procedures), and their brains may have been affected by epileptic seizures. Therefore, it is hard to generalise the findings from the studies to the general population.
Scanning Techniques
- Functional magnetic resonance imaging (fMRI): this measures changes in blood flow and oxygenation in the brain whilst the individual is performing a task. Increased blood flow suggests that the area of the brain is working harder, so it can be determined which areas of the brain are involved with particular tasks.
- Electroencephalogram (EEG): measures the electrical activity of neurons in the brain, by recording the electrical impulses that take place during synaptic transmission. The individual wears a skull cap to do this. This can detect particular patterns of activity in the brain, for example what is happening during sleep.
- Event-related potentials (ERPs): activity from an EEG recording is analysed, in order to determine the specific responses relating to a particular task. Event-related potentials are therefore the results of this- types of brainwave which are triggered by particular events.
- Post-mortem examinations: the brain is studied and analysed following death, by looking at particular areas. This is often done for individuals who have had rare disorders or dysfunction in behaviour or cognitions. Their brain will be compared to a ‘control’ brain to see if there are differences in structure.
- fMRI is low-risk, involving no radiation, and produces very detailed images. However, it is expensive, and there is a time-lag- the image taken is 5 seconds behind the initial firing of neurons (therefore, this is poor temporal resolution ). Also, the activity of individual neurons cannot be seen.
- EEGs have been very useful in diagnosing conditions such as epilepsy, and the processes involved in activities such as sleep. Brain activity can be measured almost instantaneously (a single millisecond), unlike in fMRI. However, the information gained is very generalised, so the technique can’t be used to isolate exact neural activity.
- ERPs draw on EEGs to measure more specific activity in the brain. They have excellent temporal resolution and are widely used in identifying specific behaviours and functions. Many different ERPs have been successfully identified. However, there is a lack of standardisation in the methodology used, meaning the findings are in question, and eliminating all extraneous variables in order to isolate an ERP can be difficult.
- Post-mortems have greatly enhanced medical knowledge, for example Broca and Wernicke both made use of them before neuroimaging techniques were possible. However, it is hard to establish a cause-effect link when conducting post-mortem studies. Changes in brain structure may not be related to the disorder the patient had, but due to another issue.
Circadian Rhythms
A biological rhythm is a change in the boy’s processes, in response to environmental changes. These rhythms are influenced by external and internal factors. The internal factors are the body’s internal processes- the ‘body clock’, known as endogenous pacemakers . External factors are changes in the environment, known as exogenous zeitgebers . Rhythms that last around 24 hours are known as circadian rhythms .
Sleep/wake cycle: This is an example of a circadian rhythm. The exogenous zeitgeber in this case is the daylight (or lack of it), which contributes to feelings of drowsiness or being awake. Researcher Michael Siffre studied the effect of a complete lack of daylight on his own sleep/wake cycle, by living in a cave for several months at a time. After two months in one cave, he emerged believing the date to be mid-August, but it was actually mid-September. In each experiment, his body created a natural rhythm of just beyond the usual 24 hours, and he continued to sleep and wake on a regular cycle.
Aschoff and Wever (1976) found that participants who spent 4 weeks in a bunker without natural light showed circadian rhythms of 24-25 hours, except one participant who went up to 29 hours. This suggests the natural circadian rhythm is slightly shortened by the effects of daylight. Folkard et al (1985) found that when participants were deprived of sunlight for 3 weeks, and the length of day was manipulated by the researchers to 22 hours rather than 24 (by covertly adjusting the time on the clocks), only one participant easily adjusted to the shortened day. This suggests the strength of the body’s sleep/wake cycle, as it resisted environmental changes.
- Research into circadian rhythms has useful practical applications, for example how to manage the shift patterns of night workers so that they are more productive and make fewer mistakes. This increases the usefulness of the studies.
- Research has also shown when the effects of drugs on the body are at their most and least effective. Circadian rhythms seem to have an impact on how drugs affect the body, so guidelines can be developed for patients as to when they should take drugs for maximum impact. This is another useful practical application.
- Research in this area often uses small sample sizes (only 1, in the case of Siffre), so generalisation may be difficult. Also, participants had access to artificial light, which could have acted as a confounding variable- for example, turning off a light to go to sleep may have similar effects as the end of natural light at the end of a day. The internal validity of the research is therefore in question.
Infradian & Ultradian Rhythms
Infradian rhythms.
These take place over a longer time than 24 hours. For example, the menstrual cycle . This takes place over around 28 days, although this varies between women. Hormone levels rise during the cycle, which causes the release of an egg (ovulation), then the release of the hormone progesterone which thickens the womb lining to ready the body for pregnancy. If no pregnancy occurs, the womb lining comes away, resulting in menstruation.
There is evidence that the menstrual cycle can be affected by exogenous factors. McClintock et al (1998) found that collecting pheromones (chemicals released into the air and absorbed by others, affecting their behaviour) from women with irregular cycles and rubbing them on the lips of other women caused the ‘receiver’ of the pheromones to experience changes in their cycle, bringing them closer to the pheromone ‘donor’.
Seasonal affective disorder (SAD) is another infradian rhythm, characterised by changes to mood. Sufferers feel a lowered mood, and lowered activity levels, during the winter months when daylight is shorter. As their mood changes in a predictable way through the year, this is an example of a circannual rhythm. This is thought to be caused by the hormone melatonin , which has an effect on serotonin, a neurotransmitter linked with depression. More melatonin is released during the winter, as it is released when there is a lack of daylight.
Ultradian Rhythms
These take place more than once within a 24-hour period. An example is the different stages of sleep, of which 5 distinct stages have been identified through research involving an EEG.
- Stages 1 and 2: a light sleep, where a person can be easily woken. Brain wave patterns start to slow down, becoming more ‘rhythmic’ (alpha waves) at this time.
- Stages 3 and 4: delta waves take over, which are even slower than alpha waves. This is a deep sleep; from which it is hard to wave the person- sometimes known as ’slow wave sleep’.
- Stage 5: the pons (part of the brain) paralyses the body to stop the person from ‘acting out’ their dreams. The eyelids move in a fast, jerky fashion at this point, which is correlated with dreaming. Therefore, this stage is known as REM sleep (rapid eye movement).
- Research into the menstrual cycle is likely to be affected by many variables, such as diet, stress, amount of exercise, and so on. This means that the findings of studies such as McClintock may not be valid. Other studies have found no evidence of menstrual synchrony (women’s cycles moving closer to each other’s).
- Dement and Kleitman (1957) found that REM activity was strongly correlated with dreaming. Participants woken during REM sleep were able to describe dreams in vivid detail. This supports the effect of biological rhythms on the body and brain.
- An effective treatment for SAD has been developed as a result of research in this area. Sufferers are given a light box to simulate the effects of sunlight in the dark mornings and evenings of winter, which has led to a relief in symptoms for around 60% of sufferers. This increases the practical usefulness of research into infradian rhythms.
Endogenous Pacemakers
The suprachiasmatic nucleus (SCN): This is located in the brain’s hypothalamus in both hemispheres, and is influential in the maintenance of circadian rhythms. The SCN receives information about light from a structure called the optic chiasm, which sends messages from the eye to the visual area of the cerebral cortex. This can continue even if the person’s eyes are closed, allowing the body to adjust to changing daylight patterns.
Animal studies involving the SCN have shown that if the SCN connections are destroyed, the animals no longer have a sleep/wake cycle- this was observed in chipmunks by DeCoursey et al (2000). In addition, Ralph et al (1990) found that hamsters who received SCN cells through transplant from other hamsters bred to have a 20-hour sleep/wake cycle themselves defaulted to a 20-hour sleep/wake cycle.
The pineal gland and melatonin: The SCN passes information about daylight to the pineal gland, which is located behind the hypothalamus. This gland increases melatonin production, which induces sleep and is inhibited when a person is awake. As previously seen, melatonin is a possible cause of seasonal affective disorder.
Exogenous Zeitgebers
Environmental factors have an influence on biological rhythms through a process known as ‘entrainment’. These factors work with the body’s internal processes to affect rhythms such as the sleep/wake cycle.
Light: This resets the SCN, so has a key effect on the sleep/wake cycle. Hormone production and other processes are also influenced by light. Campbell and Murphy (1998) found that skin can detect light- when participants had light shone on to the back of their knees, this affected the duration of their sleep/wake cycles- even if it was dark outside. This suggests that light is perceived not just by the eyes, and has an effect on the body.
Social cues: Infants have no set sleep/wake cycle until about 6 weeks of age, and this process generally continues until around 16 weeks, when babies are entrained. This could be due to the schedules imposed on them by parents. Similarly, the effects of jet lag can be reduced by quickly adapting to local times for sleeping and eating (not going to bed when you feel tired, for example). This suggests that the body does respond to cues in the environment.
- Damiloa et al (2000) found that other organs in the body have their own circadian rhythms in cells known as ‘peripheral oscillators’. Changing feeding patterns in mice led to changes in the rhythms of the mice’s livers, for example. This supports that there are many influences on circadian rhythms, aside from the SCN.
- Animals used in this research are often exposed to great harm, for example in the DeCoursey study many of the chipmunks were killed by predators after their sleep/wake cycle was destroyed. This raises the question of whether the research is ethically justifiable.
- Evidence suggests that exogenous zeitgebers may not actually have much of an effect on biological rhythms. Miles et al (1977) reported that a man who was blind since birth and had a sleep/wake cycle of 24.9 hours could not have his cycle adjusted by any external factors such as social cues. Instead, he had to take sedatives at night and stimulants in the morning so that he could live in the ’24-hour world’. This weakens the influence of exogenous zeitgebers on biological rhythms.
Roger Sperry’s Split Brain Experiments (1959–1968) - Image

How to cite
Articles rights and graphics.
Copyright Arizona Board of Regents Licensed as Creative Commons Attribution-NonCommercial-Share Alike 3.0 Unported (CC BY-NC-SA 3.0)
Last modified
Share this page.
IMAGES
VIDEO
COMMENTS
Conclusions. In summary, the pivotal issue in split-brain research is whether dividing the brain divides consciousness. That is, do we find evidence for the existence of one, or two conscious agents in a split-brain? Note that intermediate results may be found. Perhaps some measures indicate unified consciousness while others do not.
Conclusions. In summary, the pivotal issue in split-brain research is whether dividing the brain divides consciousness. That is, do we find evidence for the existence of one, or two conscious agents in a split-brain? Note that intermediate results may be found. Perhaps some measures indicate unified consciousness while others do not.
As the opportunities for split-brain research dwindle, Gazzaniga is busy trying to digitize the archive of recordings of tests with cohort members, some of which date back more than 50 years.
For several decades, split-brain research has provided valuable insight into the fields of psychology and neuroscience. These studies have progressed our knowledge of hemispheric specialization, language processing, the role of the corpus callosum, cognition, and even human consciousness. Following a recent empirical paper by Pinto et al ...
Lukas J. Volz, Michael S. Gazzaniga, Interaction in isolation: 50 years of insights from split-brain research, Brain, Volume 140, Issue 7, July 2017, Pages 2051-2060 ... The broad conclusions made in the article were based on meticulous observations and careful behavioural testing of these special neurological patients. As anyone with ...
A neuronal model of a global workspace in effortful cognitive tasks. Proceedings of the National Academy of Sciences of the United States of America, 95 (24), 14529-14534. Dehaene, S., & Naccache, L. (2001). Towards a cognitive neuroscience of consciousness: Basic evidence and a workspace framework.
Elizabeth Jefferies. Brain Structure and Function (2022) Forty-five years ago, Roger Sperry, Joseph Bogen and I embarked on what are now known as the modern split-brain studies. These experiments ...
The Sperry laboratory was going full tilt with experiments of all kinds on the so-called "split-brain" ( 1 ). Cats and monkeys were the main animals, and the results were clear and riveting. Train one side of the brain on a sensory task, and the other side didn't know anything about it.
Introduction. The term split-brain refers to patients in whom the corpus. " ". callosum has been cut for the alleviation of medically intrac-table epilepsy. Since the earliest reports by van Wagenen and Herren (1940) and Akelaitis (1941, 1943) on the repercus-sions of a split-brain, two narratives have emerged.
go back to reference Gazzaniga, M. S. (2014). The split-brain: Rooting consciousness in biology. Proceedings of the National Academy of Sciences of the United States of America, 111(51), 18093-18094.PubMedPubMedCentral Gazzaniga, M. S. (2014). The split-brain: Rooting consciousness in biology.
A depiction of the traditional view of the split brain syndrome (top) ... In conclusion, with two patients, and across a wide variety of tasks we have shown that severing the cortical connections between the two hemispheres does not seem to lead to two independent conscious agents within one brain. ... Forty-five years of split-brain research ...
Sperry then also studied brain patterns in frogs, cats, monkeys, and human volunteers. Sperry performed much of his research on the split-brain at California Institute of Technology, or Caltech, in Pasadena, California, where he moved in 1954. Sperry began his research on split-brain in late 1950s to determine the function of the corpus callosum.
In his new book—Tales from both sides of the brain: a life in neuroscience—Michael Gazzaniga describes a heartfelt story of his life in neuroscience in the form of a memoir. The book presents a review of Gazzaniga's work on the human split brain—the theory that the right and left hemispheres of the brain can act independently, have different strengths, and separate agendas. Gazzaniga ...
July 27, 2015. In 1939, a group of 10 people between the ages of 10 and 43, all with epilepsy, traveled to the University of Rochester Medical Center, where they would become the first people to ...
Split-brain research has been enormously influential, clarifying varieties of consciousness in the brain, and the cerebral symmetry of human motives or cognitive strategies that guide different purposes in action. ... Subsequent research has suggested that this conclusion needs some qualification. Split-brained patients can achieve some ...
The conclusion that the circuitry of the brain is fixed in early development is supported by much more evidence than I can summarize here. It has given rise to a field of research focused on "axonal guidance". ... the discovery of the duality of consciousness revealed in the split-brain experiments opened whole new fields of brain research ...
Conclusion. In two experiments, we found that automatic integration of visual information does not occur in the split-brain, yet deliberate integration remains possible. ... 50 years of insights from split-brain research. Brain 140, 2051-2060. doi: 10.1093/brain/awx139, PMID: [Google Scholar] Wahl M., Lauterbach-Soon B., Hattingen E., Jung P ...
Split-Brain Research Sperry and Gazzaniga (1967) were the first to investigate hemispheric lateralisation with the use of split-brain patients. ... Conclusion: The findings of Sperry and Gazzaniga's research highlights a number of key differences between the two hemispheres. Firstly, the left hemisphere is dominant in terms of speech and ...
Sperry (1968) - Split Brain Study. Sperry, R. W. (1968). Hemisphere deconnection and unity in conscious awareness. American Psychologist, 23 (10), 723. This is the classic biological psychology study which you will look at for your H167 AS OCR Psychology exam. You will also need this study for your OCR H567 A Level Psychology core studies exam.
Split-brain or callosal syndrome is a type of disconnection syndrome when the corpus callosum connecting the two hemispheres of the brain is severed to some degree. It is an association of symptoms produced by disruption of, or interference with, the connection between the hemispheres of the brain. The surgical operation to produce this ...
Unfortunately, much of the research into lateralisation is flawed because the split-brain procedure is rarely carried out now, meaning patients are difficult to come by. Such studies often include very few participants, and often the research takes an idiographic approach. Therefore, any conclusions drawn are representative only of those ...
Split-brain research: This involves individuals who have had surgical separation of their brain hemispheres, in order to relieve symptoms of epilepsy. Research can reveal to what extent other brain functions are lateralised. ... Sperry's research supports the conclusion that the left hemisphere is more responsible for verbal and analytical ...
Experiment Chemoaffinity Brain Function. Brain--Localization of functions Brain--Surgery Split-Brain Procedure Corpus callosum Experiments Split brain.
Study objectives: Astrocytes change their intracellular calcium (Ca2+) concentration during sleep/wakefulness states in mice. Furthermore, the Ca2+ dynamics in astrocytes vary depending on the brain region. However, whether alterations in intracellular Ca2+ concentration in astrocytes can affect sleep/wakefulness states and cortical oscillations in a brain region-dependent manner remain unclear.